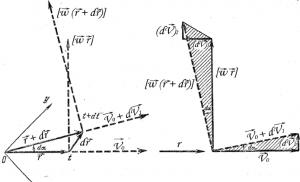
Распространение нервных импульсов. Пути передачи нервных импульсов Передача импульсов в нервной системе осуществляется посредством
В клеточной мембране располагаются Na + , K + –АТФазы, натриевые и калиевые каналы.
Na + , K + –АТФаза за счет энергии АТФ постоянно перекачивает Na + наружу и К + внутрь, создавая трансмембранный градиент концентраций этих ионов. Натриевый насос ингибируется уабаином.
Натриевые и калиевые каналы могут пропускать Na + и К + по градиентам их концентраций. Натриевые каналы блокируются новокаином, тетродотоксином, а калиевые - тетраэтиламмонием.
Работа Na + ,K + –АТФазы, натриевых и калиевых каналов может создавать на мембране потенциал покоя и потенциал действия.
Потенциал покоя – это разность потенциалов между наружной и внутренней мембраной в условиях покоя, когда натриевые и калиевые каналы закрыты. Его величина составляет -70мВ, он создается в основном концентрацией K + и зависит от Na + и Cl - . Концентрация К + внутри клетки составляет 150 ммоль/л, снаружи 4-5 ммоль/л. Концентрация Na + внутри клетки составляет 14 ммоль/л, снаружи 140 ммоль/л. Отрицательный заряд внутри клетки создают анионы (глутамат, аспартат, фосфаты), для которых клеточная мембрана непроницаема. Потенциал покоя одинаков на всем протяжении волокна и не является специфической особенностью нервных клеток.
Раздражение нерва может приводит к возникновению потенциала действия.
Потенциал действия – это кратковременное изменение разности потенциала между наружной и внутренней мембраной в момент возбуждения. Потенциал действия зависит от концентрации Na + и возникает по принципу «все или ничего».
Потенциал действия состоит из следующих стадий:
1. Локальный ответ . Если при действии стимула происходит изменение потенциала покоя до пороговой величины -50мВ, то открываются натриевые каналы, имеющие более высокую пропускную способность, чем калиевые.
2. Стадия деполяризации. Поток Na + внутрь клетки приводит сначала к деполяризации мембраны до 0 мВ, а затем к инверсии полярности до +50мВ.
3. Стадия реполяризации. Натриевые каналы закрываются, а калиевые открываются. Выход К + из клетки восстанавливает мембранный потенциал до уровня потенциала покоя.
Ионные каналы открываются на непродолжительное время и после их закрытия натриевый насос восстанавливает исходное распределение ионов по сторонам мембраны.
Нервный импульс
В отличие от потенциала покоя, потенциал действия охватывает лишь очень небольшой участок аксона (в миелинизированных волокнах – от одного перехвата Ранвье до соседнего). Возникнув в одном участке аксона, потенциал действия вследствие диффузии ионов из этого участка вдоль волокна снижает потенциал покоя в соседнем участке и вызывает здесь то же развитие потенциала действия. Благодаря этому механизму потенциал действия распространяется по нервным волокнам и называется нервным импульсом .
В миелинизированном нервном волокне натриевые и калиевые ионные каналы расположены в немиелинизированных участках перехватов Ранвье, где мембрана аксона контактирует с межклеточной жидкостью. Вследствие этого нервный импульс перемещается «скачками»: ионы Na + , поступающие внутрь аксона при открытии каналов в одном перехвате, диффундируют вдоль аксона по градиенту потенциалов до следующего перехвата, снижают здесь потенциал до пороговых значений и тем самым индуцируют потенциал действия. Благодаря такому устройству скорость поведения импульса в миелинизированном волокне в 5-6 раз больше, чем в немиелинизированных волокнах, где ионные каналы расположены равномерно по всей длине волокна и потенциал действия перемещается не скачками, а плавно.
Синапс: виды, строение и функции
Вальдаер в 1891г. сформулировал нейронную теорию , согласно которой нервная система состоит из множества отдельных клеток – нейронов. В ней оставался неясным вопрос: каков механизм коммуникации между единичными нейронами? Ч. Шеррингтон в 1887г. для объяснения механизма взаимодействия нейронов ввел термин «синапс» и «синаптическая передача».
Кандидат биологических наук Л. Чайлахян, научный сотрудник Института биофизики АН СССР
Читательница журнала Л. Горбунова (деревня Цыбино, Московской области) пишет нам: «Меня интересует механизм, передачи сигналов по нервным, клеткам».
Лауреаты Нобелевской премии 1963 года (слева направо): А. Ходжкин, Э. Хаксли, Д. Экклс.
Представления ученых о механизме передачи нервного импульса претерпели в последнее время существенное изменение. До недавнего времени в науке господствовали взгляды Бернштейна.
Мозг человека, без сомнения, высшее достижение природы. В килограмме нервной ткани заключена квинтэссенция всего человека, начиная от регуляции жизненных функций - работы сердца, легких, пищеварительного тракта, печени - и кончая его духовным миром. Здесь - наши мыслительные способности, всё наше мироощущение, память, разум, наше самосознание, наше «я». Познание механизмов работы мозга - это познание самого себя.
Велика и заманчива цель, но неимоверно сложен объект исследования. Шутка сказать, этот килограмм ткани представляет собой сложнейшую систему связи десятков миллиардов нервных клеток.
Однако первый существенный шаг к познанию работы мозга уже сделан. Может быть, он один из самых легких, но он чрезвычайно важен для всего дальнейшего.
Я имею в виду исследование механизма передачи нервных импульсов - сигналов, бегущих по нервам, как по проводам. Именно эти сигналы являются той азбукой мозга, с помощью которой органы чувств посылают в центральную нервную систему сведения-депеши о событиях во внешнем мире. Нервными импульсами зашифровывает мозг свои приказы мышцам и различным внутренним органам. Наконец, на языке этих сигналов говорят между собой отдельные нервные клетки и нервные центры.
Нервные клетки - основной элемент мозга - разнообразны по величине, по форме, но в принципе обладают единым строением. Каждая нервная клетка состоит из трех частей: из тела, длинного нервного волокна - аксона (длина его у человека от нескольких миллиметров до метра) и нескольких коротких ветвистых отростков - дендритов. Нервные клетки изолированы друг от друга оболочками. Но все же клетки взаимодействуют между собой. Происходит это в месте стыка клеток; этот стык называется «синапс». В синапсе встречаются аксон одной нервной клетки и тело или дендрит другой клетки. Причем интересно, что возбуждение может передаваться только и одном направлении: от аксона к телу или дендриту, но ни в коем случае не обратно. Синапс - это как бы кенотрон: он пропускает сигналы только в одном направлении.
В проблеме изучения механизма нервного импульса и его распространения можно выделить два основных вопроса: природа проведения нервного импульса или возбуждения в пределах одной клетки - по волокну и механизм передачи нервного импульса от клетки к клетке - через синапсы.
Какова природа сигналов, передающихся от клетки к клетка по нервным волокнам?
Этой проблемой человек интересовался уже давно, Декарт предполагал, что распространение сигнала связано с переливанием жидкости по нервам, как по трубкам. Ньютон думал, что это чисто механический процесс. Когда появилась электромагнитная теория, ученые решили, что нервный импульс аналогичен движению тока по проводнику со скоростью, близкой к скорости распространения электромагнитных колебаний. Наконец, с развитием биохимии появилась точка зрения, что движение нервного импульса - это распространение вдоль по нервному волокну особой биохимической реакции.
И всё же ни одно из этих представлений не оправдалось.
В настоящее время природа нервного импульса раскрыта: это удивительно тонкий электрохимический процесс, в основе которого лежит перемещение ионов через оболочку клетки.
Большой вклад в раскрытие этой природы внесли работы трех ученых: Алана Ходжкина, профессора биофизики Кембриджского университета; Эндрью Хаксли, профессора физиологии Лондонского университета, и Джона Экклса, профессора физиологии австралийского университета в Канберре. Им присуждена Нобелевская премия в области медицины за 1963 год,
Впервые предположение об электрохимической природе нервного импульса высказал известный немецкий физиолог Бернштейн в начале нашего столетия.
К началу двадцатого века было довольно многое известно о нервном возбуждении. Ученые уже знали, что нервное волокно можно возбудить электрическим током, причем возбуждение всегда возникает под катодом - под минусом. Было известно, что возбужденная область нерва заряжается отрицательно по отношению к невозбужденному участку. Было установлено, что нервный импульс в каждой точке длится всего 0,001-0,002 секунды, что величина возбуждения не зависит от силы раздражения, как громкость звонка в нашей квартире не зависит от того, как сильно мы нажимаем на кнопку. Наконец, ученые установили, что носителями электрического тока в живых тканях являются ионы; причем внутри клетки основной электролит - соли калия, а в тканевой жидкости - соли натрия. Внутри большинства клеток концентрация ионов калия в 30-50 раз больше, чем в крови и в межклеточной жидкости, омывающей клетки.
И вот на основании всех этих данных Бернштейн предположил, что оболочка нервных и мышечных клеток представляет собой особую полупроницаемую мембрану. Она проницаема только для ионов К + ; для всех остальных ионов, в том числе и для находящихся внутри клетки отрицательно заряженных анионов, путь закрыт. Ясно, что калий по законам диффузии будет стремиться выйти из клетки, в клетке возникает избыток анионов, и по обе стороны мембраны появится разность потенциалов: снаружи - плюс (избыток катионов), внутри - минус (избыток анионов). Эта разность потенциалов получила название потенциала покоя. Таким образом, в покое, в невозбужденном состоянии внутренняя часть клетки всегда заряжена отрицательно по сравнению с наружным раствором.
Бернштейн предположил, что в момент возбуждения нервного волокна происходят структурные изменения поверхностной мембраны, ее поры как бы увеличиваются, и она становится проницаемой для всех ионов. При этом, естественно, разность потенциалов исчезает. Это и вызывает нервный сигнал.
Мембранная теория Бернштейма быстро завоевала признание и просуществовала свыше 40 лет, вплоть до середины нашего столетия.
Но уже в конце 30-х годов теория Бернштейна встретилась с непреодолимыми противоречиями. Сильный удар ей был нанесен в 1939 году тонкими экспериментами Ходжкина и Хаксли. Эти ученые впервые измерили абсолютные величины мембранного потенциала нервного волокна в покое и при возбуждении. Оказалось, что при возбуждении мембранный потенциал не просто уменьшался до нуля, а переходил через ноль на несколько десятков милливольт. То есть внутренняя часть волокна из отрицательной становилась положительной.
Но мало ниспровергнуть теорию, надо заменить ее другой: наука не терпит вакуума. И Ходжкин, Хаксли, Катц в 1949-1953 годах предлагают новую теорию. Она получает название натриевой.
Здесь читатель вправе удивиться: до сих пор о натрии не было речи. В этом все и дело. Ученые установили с помощью меченых атомов, что в передаче нервного импульса замешаны не только ионы калия и анионы, но и ионы натрия и хлора.
В организме достаточно ионов натрия и хлора, все знают, что кровь соленая на вкус. Причем натрия в межклеточной жидкости в 5-10 раз больше, чем внутри нервного волокна.
Что же это может означать? Ученые предположили, что при возбуждении в первый момент резко увеличивается проницаемость мембраны только для натрия. Проницаемость становится в десятки раз больше, чем для ионов калия. А так как натрия снаружи в 5-10 рез больше, чем внутри, то он будет стремиться войти в нервное волокно. И тогда внутренняя часть волокна станет положительной.
А через какое-то время - после возбуждения - равновесие восстанавливается: мембрана начинает пропускать и ионы калия. И они выходят наружу. Тем самым они компенсируют тот положительный заряд, который был внесен внутрь волокна ионами натрия.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно непонятно, каким образом больший по размеру ион проходит там, где не может пройти меньший.
Нужно было решительно изменить взгляд на механизм перехода ионов через мембраны. Ясно, что только рассуждениями о порах в мембране здесь не обойтись. И тогда была высказана идея, что ионы могут пересекать мембрану совершенно другим способом, с помощью тайных до поры до времени союзников - особых органических молекул-переносчиков, спрятанных в самой мембране. С помощью такой молекулы ионы могут пересекать мембрану в любом месте, а не только через поры. Причем эти молекулы-такси хорошо различают своих пассажиров, они не путают ионы натрия с ионами калия.
Тогда общая картина распространения нервного импульса будет иметь следующий вид. В покое молекулы-переносчики, заряженные отрицательно, мембранным потенциалом прижаты к наружной границе мембраны. Поэтому проницаемость для натрия очень мала: в 10-20 раз меньше, чем для ионов калия. Калий может пересекать мембрану через поры. При приближении волны возбуждения уменьшается давление электрического поля на молекулы-переносчики; они сбрасывают свои электростатические «оковы» и начинают переносить ионы натрия внутрь клетки. Это еще больше уменьшает мембранный потенциал. Идет как бы цепной процесс перезарядки мембраны. И этот процесс непрерывно распространяется вдоль нервного волокна.
Интересно, что нервные волокна тратят на свою основную работу - проведение нервных импульсов - всего около 15 минут в сутки. Однако готовы к этому волокна в любую секунду: все элементы нервного волокна работают без перерыва - 24 часа в сутки. Нервные волокна в этом смысле подобны самолетам-перехватчикам, у которых непрерывно работают моторы для мгновенного вылета, однако сам вылет может состояться лишь раз в несколько месяцев.
Мы познакомились сейчас с первой половиной таинственного акта прохождения нервного импульса - вдоль одного волокна. А как же передается возбуждение от клетки к клетке, через места стыков - синапсы. Этот вопрос был исследован в блестящих опытах третьего нобелевского лауреата, Джона Экклса.
Возбуждение не может непосредственно перейти с нервных окончаний одной клетки на тело или дендриты другой клетки. Практически весь ток вытекает через синаптическую щель в наружную жидкость, и в соседнюю клетку через синапс попадает ничтожная его доля, неспособная вызвать возбуждение. Таким образом, в области синапсов электрическая непрерывность в распространении нервного импульса нарушается. Здесь, на стыке двух клеток, в силу вступает совершенно другой механизм.
Когда возбуждение подходит к окончанию клетки, к месту синапса, в межклеточную жидкость выделяются физиологически активные вещества - медиаторы, или посредники. Они становятся связующим звеном в передаче информации от клетки к клетке. Медиатор химически взаимодействует со второй нервной клеткой, изменяет ионную проницаемость ее мембраны - как бы пробивает брешь, в которую устремляются многие ионы, в том числе и ионы натрия.
Итак, благодаря работам Ходжкина, Хаксли и Экклса важнейшие состояния нервной клетки - возбуждение и торможение - можно описать в терминах ионных процессов, в терминах структурно-химических перестроек поверхностных мембран. На основании этих работ уже можно делать предположения о возможных механизмах кратковременной и долговременной памяти, о пластических свойствах нервной ткани. Однако это разговор о механизмах в пределах одной или нескольких клеток. Это лишь, азбука мозга. По-видимому, следующий этап, возможно, гораздо более трудный, - вскрытие законов, по которым строится координирующая деятельность тысяч нервных клеток, распознание языка, на котором говорят между собой нервные центры.
Мы сейчас в познании работы мозга находимся на уровне ребенка, который узнал буквы алфавита, но не умеет связывать их в слова. Однако недалеко время, когда ученые с помощью кода - элементарных биохимических актов, происходящих в нервной клетке, прочтут увлекательнейший диалог между нервными центрами мозга.
Детальное описание иллюстраций
Представления ученых о механизме передачи нервного импульса претерпели в последнее время существенное изменение. До недавнего времени в науке господствовали взгляды Бернштейна. По его мнению, в состоянии покоя (1) нервное волокно заряжено положительно снаружи и отрицательно внутри. Это объяснялось тем, что сквозь поры в стенке волокна могут проходить только положительно заряженные ионы калия (К +); большие по размерам отрицательно наряженные анионы (А –) вынуждены оставаться внутри и создавать избыток отрицательных зарядов. Возбуждение (3) по Бернштейну сводится к исчезновению разности потенциалов, которое вызывается тем, что размер пор увеличивается, анионы выходят наружу и выравнивают ионный баланс: количество положительных ионов становится равным количеству отрицательных. Работа лауреатов Нобелевской премии 1963 года А. Ходжкпна, Э. Хаксли и Д. Экклса изменила наши прежние представления. Доказано, что в нервном возбуждении участвуют также положительные ионы натрия (Na +), отрицательные ноны хлора (Сl –) и отрицательно заряженные молекулы-переносчики. Покоящееся состояние (3) образуется в принципе так же, как и считалось раньше: избыток положительных ионов - снаружи нервного волокна, избыток отрицательных - внутри. Однако установлено, что при возбуждении (4) происходит не выравнивание зарядов, а перезарядка: снаружи образуется избыток отрицательных ионов, а внутри - избыток положительных. Объясняется это тем, что при возбуждении молекулы-переносчики начинают перевозить сквозь стенку положительные ионы натрия. Таким образом, нервный импульс (5) - это перемещающаяся вдоль волокна перезарядка двойного электрического слоя. А от клетки к клетке возбуждение передается своеобразным химическим «тараном» (6) - молекулой ацетилхолина, которая помогает ионам прорываться сквозь стенку соседнего нервного волокна.
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны . Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами , служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия . При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом . Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими ) пузырьками , в каждом из которых находятся особые соединения — нейромедиаторы . При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом ). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Раздражения рецепторов вызывают преобразование воздействующей энергии раздражителя в нервные импульсы , передача которых в нервной системе осуществляется с помощью синапсов .
Функциональные структуры клеточной мембраны. Клеточная мембрана (оболочка клетки) представляет собой тонкую липопротеиновую пластинку, содержание липидов – около 40%, белков – 60%. Схематично клеточную мембрану можно представить следующим образом: мембрана состоит из двойного слоя молекул фосфолипидов, покрытого изнутри слоем белковых молекул, а снаружи – слоем молекул сложных углеводов. В клеточной мембране имеются тончайшие канальцы – ионные каналы, обладающие селективностью. Различают каналы, которые пропускают только один ион (натрия, калия, кальция, хлора), либо несколько.
Потенциал покоя и потенциал действия. В состоянии покоя в протоплазме нервной клетки концентрация ионов калия более, чем в 30 раз превышает концентрацию этих ионов в наружном растворе. Мембрана практически непроницаема для натрия, тогда как калий проходит сквозь нее. Диффузия ионов калия из протоплазмы во внешнюю жидкость очень высокая, что и придает наружной мембране положительный заряд, а внутренней – отрицательный. Таким образом, концентрация ионов калия является основным фактором, формирующим и определяющим величину потенциала покоя (ПП).
При действии на клетку раздражения проницаемость мембраны для ионов натрия резко повышается и становится, примерно, в 10 раз больше проницаемости для ионов калия. Поэтому поток положительно заряженных ионов калия из протоплазмы во внешний раствор уменьшается, а возрастает поток положительно заряженных ионов натрия из внешнего раствора в протоплазму клетки. Это приводит к перезарядке мембраны, наружная поверхность становится заряженной электро-отрицательно, а внутренняя - положительно (фаза деполяризации).
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость для ионов натрия снова понижается, а проницаемость ее для ионов калия возрастает. И в итоге этих двух процессов снова наружная мембрана приобретает положительный заряд, а внутренняя – отрицательный (фаза реполяризации).
Мгновенное увеличение проницаемости для ионов натрия и их проникновение в клетку достаточно, чтобы изменить знак мембранного потенциала и возникает потенциал действия (ПД), который распространяется по аксону с довольно большой скоростью, длительность ПД обычно составляет 1-3 мс.
Синаптическая передача информации. Место передачи возбуждения с одного нейрона на другой называется синапсом (в переводе с греческого – контакт). Синапс представляет собой мембраны двух соседних нейронов (пресинаптическая и постсинаптическая мембраны) и пространство между ними, которое называется синаптической щелью.
Различают аксо-соматические синапсы, сформированные мембранами аксона и телом (сомой) другого нейрона, аксо-дендритные, состоящие из мембраны аксона и дендритами другого нейрона, аксо-аксональные, при которых аксон подходит к аксону другого нейрона. Синапс между аксонами и мышечными волокнами называется нейромышечной пластинкой.
Нервный импульс по аксону достигает окончания аксона и вызывает открытие каналов для кальция на пресинаптической мембране. Здесь, на пресинаптической мембране находятся везикулы (пузырьки), которые содержат биологически активные вещества – медиаторы.
Открытие кальцевых каналов приводит к деполяризации на пресинаптической мембране. Кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. Затем пузырьки лопаются и все содержимое поступает в синаптическую щель. Далее молекулы медиатора связываются со специальными белковыми молекулами (рецепторами) , которые находятся на мембране другого нейрона – на постсинаптической мембране.
Когда молекулы медиаторов связываются с рецепторами, то на постсинаптической мембране открываются каналы для ионов натрия и калия, вызывая на ней изменение потенциала (деполяризацию). Этот потенциал получил название – постсинаптический потенциал (ПСП). В зависимости от характера открытых ионных каналов возникает возбудительный (ВПСП) или тормозный (ТПСП) постсинаптические потенциалы
Таким образом, возбуждение (ПД) нейрона в синапсе превращается из электрического импульса в химический импульс (выброс медиатора из везикул).
Время между началом пресинаптической деполяризации и постсинаптической реакцией составляет 0,5 мс, это – синаптическая задержка.
Основные медиаторы: ацетилхолин, моноамины (серотонин, гистамин), катехоламины (дофамин, норадреналин, адреналин), аминокислоты (глутамат, глицин, аспартат, гамма-аминомаслянная кислота – ГАМК, аланин), пептиды, вазопрессин, окситоцин, аденозин, АТФ и др.
Спинной мозг
Спинной мозг , по внешнему виду представляет собой длинный, цилиндрической формы, уплощенный спереди назад тяж. В связи с этим поперечный диаметр спинного мозга больше переднезаднего.
Спинной мозг располагается в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В этом месте из спинного мозга (верхняя его граница) выходят корешки, образующие правый и левый спинномозговые нервы. Нижняя граница спинного мозга соответствует уровню 1-11 поясничных позвонков. Ниже этого уровня верхушка мозгового конуса спинного мозга продолжается в тонкую терминальную (концевую) нить. Терминальная нить в своих верхних отделах еще содержит нервную ткань и представляет собой рудимент каудального конца спинного мозга. Эта часть терминальной нити, получившая название внутренней, окружена корешками поясничных и крестцовых спинномозговых нервов и вместе с ними находится в слепо заканчивающемся мешке, образованном твердой оболочкой спинного мозга. У взрослого человека внутренняя часть терминальной нити имеет длину около 15 см. Ниже уровня 2 крестцового позвонка терминальная нить представляет собой соединительнотканное образование, являющееся продолжением всех трех оболочек спинного мозга и получившее название наружной части терминальной нити. Длина этой части около 8 см. Заканчивается она на уровне тела 2 копчикового позвонка, срастаясь с его надкостницей.
Длина спинного мозга у взрослого человека в среднем 43 см (у мужчин 45 см, у женщин 41-42 см), масса - около 34-38 г, что составляет примерно 2 % от массы головного мозга.
В шейном и поясничнокрестцовом отделах спинного мозга обнаруживаются два заметных утолщения: шейное утолщение и пояснично-крестцовое утолщение. Образование утолщений объясняется тем, что от шейного и поясничнорестцового отделов спинного мозга осуществляется иннервация соответственно верхних и нижних конечностей. В этих отделах в спинном мозге имеется большее по сравнению с другими отделами количество нервных клеток и волокон. На нижних отделах спинной мозг постепенно суживается и образует мозговой конус.
На передней поверхности спинного мозга видна передняя срединная щель, которая вдается в ткань спинного мозга глубже, чем задняя срединная борода. Эти борозды являются границами, разделяющими спинной мозг на две симметричные половины. В глубине задней срединной борозды имеется проникающая почти во всю толщу белого вещества глиальная задняя срединная перегородка. Эта перегородка доходит до задней поверхности серого вещества спинного мозга.
На передней поверхности спинного мозга, с каждой стороны от передней щели, проходит переднелатеральная борода. Она является местом выхода из спинного мозга передних (двигательных) корешков спинномозговых нервов и границей на поверхности спинного мозга между передним и боковым канатиками. На задней поверхности на каждой половине спинного мозга имеется заднелатеральная борозда, место проникновения в спинной мозг задних чувствительных корешков спинномозговых нервов. Эта борозда служит границей между боковым и задним канатиками.
Передний корелток состоит ив отростков двигательных (моторных) нервных клеток, расположенных в переднем роге серого вещества спинного мозга. Задний корешок - чувствительный, представлен совокупностью проникающих в спинной мозг центральных отростков псевдоуниполярных клеток, тела которых образуют спинномозговой узел, лежащий у места соединения заднего корешка с передним. На всем протяжении спинного мозга с каждой его стороны отходит 31 пара корешков. Передний и задний корешки у внутреннего края межпозвоночного отверстия сближаются, сливаются друг с другом и образуют спинномозговой нерв. Таким образом, ив корешков образуется 31 пара спинномозговых нервов. Участок спинного мозга, соответствующий двум парам корешков (два передних и два задних), называют сегментон.
Для врача очень важно знать топографические взаимоотношения сегментов спинного мозга с позвоночным столбом (скелетотопия сегментов) . Протяженность спинного мозга значительно меньше длины позвоночного столба, поэтому порядковый номер какого-либо сегмента спинного мозга и уровень его положения, начиная с нижнего шейного отдела, не соответствует порядковому номеру одноименного позвонка. Положение сегментов по отношению к позвонкам можно определить следующим образом. Верхние шейные сегменты расположены на уровне соответствующих их порядковому номеру тел позвонков. Нижние шейные и верхние грудные сегменты лежат на один по- звонок выше, чем тела соответствующих позвонков. В среднем грудном отделе эта разница между соответствующим сегментом спинного мозга и телом позвонка увеличивается уже на 2 позвонка, в нижнем грудном - на З. Поясничные сегменты спинного мозга лежат в позвоночном канале на уровне тел 10, 11 грудных позвонков, крестцовые и копчиковый сегменты - на уровне 12 грудного и 1 поясничного позвонков.
Спинной мозг состоит из нервных клеток и волокон серого вещества, имеющего на поперечном срезе вид буквы В или бабочки с расправленными крыльями. За периферии от серого вещества находится белое вещество, образованное только нервными волокнами.
В сером веществе спинного мозга имеется центральный канал. Он является остатком полости нервной трубки и содержит спинномозговую жидкость. Верхний конец канала сообщается с 9 желудочком, а нижний, несколько расширяясь, образует слепо заканчивающийся концевой желудочек. Стенки центрального канала спинного мозга выстланы эпендимой, вокруг которой находится центральное студенистое (серое) вещество. У взрослого человека центральный канал в различных отделах спинного мозга, а иногда и на всем протяжении зарастает.
Серое вещество, на протяжении спинного мозга справа и слева от центрального канала образует симметричные серые столбы. Кпереди и кзади от центрального канала спинного мозга эти серые столбы связаны друг с другом тонкими пластинками серого вещества, получившими название передней и задней спаек.
В каждом столбе серого вещества различают переднюю его часть - передний столб и заднюю часть - задний столб. За уровне нижнего шейного, всех грудных и двух верхних поясничных сегментов спинного мозга.
Серое вещество с каждой стороны образует боковое выпячивание - боковой столб. В других отделах спинного мозга (выше 8 шейного и ниже 2 поясничного сегментов) боковые столбы отсутствуют.
За поперечном срезе спинного мозга столбы серого вещества с каждой стороны имеют вид рогов. Выделяют более широкий передний рог и узкий задний рог1, соответствующие переднему и заднему столбам. Боковой рог, соответствует боковому промежуточному столбу (автономному) серого вещества.
В передних рогах расположены крупные нервные корешковые клетки - двигательные (эфферентные) нейроны. Эти нейроны образуют 5 ядер: два латеральных (передне и заднелатеральное), два медиальных (передне и заднемедиальное) и центральное ядро. Задние рога спинного мозга представлены преимущественно более мелкими клетками. В составе задних, или чувствительных, корешков находятся центральные отростки псевдоуниполярных клеток, расположенных в спинномозговых (чувствительных) узлах.
Серое вещество задних рогов спинного мозга неоднородно. Основная масса нервных клеток заднего рога образует собственное его ядро. В белом веществе, непосредственно примыкающем к верхушке заднего рога серого вещества, выделяют пограничную зону. Кпереди от последней в сером веществе расположена губчатая зона, которая получила свое название в связи с наличием в этом отделе крупнопетлистой глиальной сети, содержащей нервные клетки. Еще более кпереди выделяется студенистое вещество, состоящее из мелких нервных клеток. Отростки нервных клеток студенистого вещества, губчатой зоны и диффузно рассеянных во всем сером веществе пучковых клеток осуществляют связь с несколькими соседними сегментами. Как правило, они заканчиваются синапсами с нейронами, расположенными в передних рогах своего сегмента, а также выше и нижележащих сегментов. Направляясь от задних рогов серого вещества к передним рогам, отростки этих клеток располагаются по периферии серого вещества, образуя возле него узкую каемку белого вещества. Эти пучки нервных волокон получили название передних, латеральных и задних собственных пучков. Клетки всех ядер задних рогов серого вещества - это, как правило, вставочные (промежуточные, или кондукторные) нейроны. Нейриты, отходящие от нервных клеток, совокупность которых составляет центральное и грудное ядра задних рогов, направляются в белом веществе спинного мозга к головному мозгу.
Промежуточная зона серого вещества спинного мозга расположена между передним и задним рогами. Здесь на протяжении с 8 шейного по 2 поясничный сегмент имеется выступ серого вещества - боковой рог.
В медиальной части основания бокового рога заметно хорошо очерченное прослойкой белого вещества трудное ядро, состоящее из крупных нервных клеток. Это ядро тянется вдоль всего заднего столба серого вещества в виде клеточного тяжа (ядро Кларка) . Наибольший диаметр этого ядра на уровне от 11 грудного до 1 поясничного сегмента. В боковых рогах находятся центры симпатической части вегетативной нервной системы в виде нескольких групп мелких нервных клеток, объединенных в латеральное промежуточное (серое) вещество. Аксоны этих клеток проходят через передний рог и выходят из спинного мозга в составе передних корешков.
В промежуточной зоне расположено центральное промежуточное (серое) вещество, отростки клеток которого участвуют в образовании спиномозжечкового пути. На уровне шейных сегментов спинного мозга между передним и задним рогами, а на уровне верхнегрудных сегментов - между боковыми и задним рогами в белом веществе, примыкающем к серому, расположена ретикулярная формация. Ретикулярная формация имеет здесь вид тонких перекладин серого вещества, пересекающихся в различных направлениях, и состоит из нервных клеток с большим количеством отростков.
Серое вещество спинного мозга с задними и передними корешками спинномозговых нервов и собственными пучками белого вещества, окаймляющими серое вещество, образует собственный, или сегментарный, аппарат спинного мозга. Основное назначение сегментарного аппарата как филогенетически наиболее старой части спинного мозга - осуществление врожденных реакций (рефлексов) в ответ на раздражение (внутреннее или внешнее). И. П. Павлов определил этот вид деятельности сегментарного аппарата спинного мозга термином "безусловные рефлексы".
Белое вещество, как отмечалось, локализуется кнаружи от серого вещества. Борозды спинного мозга разделяют белое вещество на симметрично расположенные справа и слева три канатика. Передний канатик, находится между передней срединной щелью и передней латеральной бороздой. Н белом веществе кзади от передней срединной щели различают переднюю белую спайку, которая соединяет передние канатики правой и левой сторон. Задний канатик, находится между задней срединной и задней латеральной бороздами. Боковой канатик- это участок белого вещества между передней и задней латеральными бороздами.