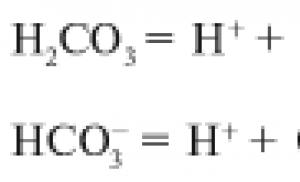Cum diferă transportul activ de transportul pasiv? Transportul activ al ionilor și moleculelor pe o membrană. Transport activ de substanțe
Transport pasiv include difuzie simplă și facilitată – procese care nu necesită energie. Difuzia este transportul moleculelor și ionilor printr-o membrană dintr-o zonă de mare într-o zonă de concentrație scăzută, adică substanțele curg de-a lungul unui gradient de concentrație. Difuzia apei prin membranele semipermeabile se numește osmoză. Apa este, de asemenea, capabilă să treacă prin porii membranei formați din proteine și molecule de transport și ioni de substanțe dizolvate în ea.Mecanismul de difuzie simplă realizează transferul de molecule mici (de exemplu, O2, H2O, CO2); acest proces are specificitate scăzută și are loc cu o rată proporțională cu gradientul de concentrație al moleculelor transportate pe ambele părți ale membranei. Difuzia facilitată are loc prin canale și/sau proteine purtătoare care au specificitate pentru moleculele transportate. Proteinele transmembranare acționează ca canale ionice, formând pori mici de apă prin care moleculele și ionii solubili în apă sunt transportați de-a lungul unui gradient electrochimic. Proteinele transportoare sunt, de asemenea, proteine transmembranare care suferă modificări conformaționale reversibile care permit transportul unor molecule specifice prin plasmalemă. Ele funcționează atât în mecanismele de transport pasiv, cât și în cele active.
Transport activ este un proces consumator de energie prin care transportul moleculelor se realizează folosind proteine purtătoare împotriva unui gradient electrochimic. Un exemplu de mecanism care asigură transportul activ direcționat opus al ionilor este pompa de sodiu-potasiu (reprezentată de proteina purtătoare Na+-K+-ATPaza), datorită căreia ionii Na+ sunt îndepărtați din citoplasmă, iar ionii K+ sunt transferați simultan în aceasta. Concentrația de K+ în interiorul celulei este de 10-20 de ori mai mare decât în exterior, iar concentrația de Na este invers. Această diferență în concentrațiile ionilor este asigurată de activitatea pompei (Na*-K*>. Pentru a menține această concentrație, trei ioni de Na sunt transferați din celulă pentru fiecare doi ioni K* în celulă. O proteină din membrană ia parte în acest proces, îndeplinind funcția unei enzime care descompune ATP, eliberând energia necesară pentru a funcționa pompa.
Participarea proteinelor membranare specifice în transportul pasiv și activ indică specificitatea ridicată a acestui proces. Acest mecanism asigură menținerea constantă a volumului celular (prin reglarea presiunii osmotice), precum și a potențialului membranar. Transportul activ al glucozei în celulă este realizat de o proteină purtătoare și este combinat cu transferul unidirecțional al ionului Na+.
Transport usor fluxul ionic este mediat de proteine transmembranare speciale - canale ionice care asigură transportul selectiv al anumitor ioni. Aceste canale constau din sistemul de transport în sine și un mecanism de deschidere care deschide canalul pentru o perioadă de timp ca răspuns la (a) o modificare a potențialului membranei, (b) stresul mecanic (de exemplu, în celulele părului urechii interne), (c) legarea unui ligand (molecula semnal sau ion).
Transportul moleculelor mici prin membrană.
Transportul pe membrană poate implica transportul unidirecțional al moleculelor unei substanțe sau transportul în comun a două molecule diferite în direcții identice sau opuse.
Prin ea trec diferite molecule cu viteze diferite și, cu cât dimensiunea moleculelor este mai mare, cu atât viteza de trecere a acestora prin membrană este mai mică. Această proprietate definește membrana plasmatică ca o barieră osmotică. Apa și gazele dizolvate în ea au capacitatea maximă de penetrare. Una dintre cele mai importante proprietăți ale membranei plasmatice este asociată cu capacitatea de a trece diferite substanțe în sau din celulă. Acest lucru este necesar pentru a menține constanța compoziției sale (adică homeostazia).
Transport de ioni.
Spre deosebire de membranele cu două straturi lipidice artificiale, membranele naturale și, în primul rând, membrana plasmatică, sunt încă capabile să transporte ioni. Permeabilitatea pentru ioni este scăzută, iar viteza de trecere a diferiților ioni nu este aceeași. Rată de trecere mai mare pentru cationi (K+, Na+) și mult mai mică pentru anioni (Cl-). Transportul ionilor prin plasmalemă are loc datorită participării proteinelor de transport membranare - permeazele - în acest proces. Aceste proteine pot transporta o substanță într-o direcție (uniport) sau mai multe substanțe simultan (symport), sau, împreună cu importul unei substanțe, pot elimina alta din celulă (antiport). De exemplu, glucoza poate pătrunde în celule simportal împreună cu ionul Na+. Transportul ionilor poate avea loc de-a lungul unui gradient de concentrație- pasiv fără consum suplimentar de energie. De exemplu, ionul Na+ pătrunde în celulă din mediul extern, unde concentrația sa este mai mare decât în citoplasmă.
Prezența canalelor și purtătorilor de proteine ar părea să conducă la o echilibrare a concentrațiilor de ioni și substanțe cu greutate moleculară mică de pe ambele părți ale membranei. De fapt, nu este așa: concentrația de ioni din citoplasma celulelor diferă mult nu numai de cea din mediul extern, ci chiar și de plasma sanguină care spală celulele din corpul animalelor.
Se dovedește că în citoplasmă concentrația de K+ este de aproape 50 de ori mai mare, iar Na+ este mai mică decât în plasma sanguină. Mai mult, această diferență se menține numai într-o celulă vie: dacă celula este ucisă sau procesele metabolice din ea sunt suprimate, atunci după un timp diferențele ionice de pe ambele părți ale membranei plasmatice vor dispărea. Pur și simplu puteți răci celulele la +20C, iar după un timp concentrațiile de K+ și Na+ de pe ambele părți ale membranei vor deveni aceleași. Când celulele sunt încălzite, această diferență este restabilită. Acest fenomen se datorează faptului că în celule există purtători de proteine membranare care lucrează împotriva gradientului de concentrație, consumând în același timp energie datorită hidrolizei ATP. Acest tip de muncă se numește transport activ, și se realizează cu ajutorul proteinelor pompe ionice. Membrana plasmatică conține o moleculă de pompă cu două subunități (K+ + Na+), care este, de asemenea, o ATPază. În timpul funcționării, această pompă pompează 3 ioni Na+ într-un ciclu și pompează 2 ioni K+ în celulă împotriva gradientului de concentrație. În acest caz, o moleculă de ATP este cheltuită pentru fosforilarea ATPazei, în urma căreia Na+ este transferat prin membrană de la celulă, iar K+ este capabil să intre în contact cu molecula de proteină și apoi este transportat în celulă. Ca urmare a transportului activ cu ajutorul pompelor membranare, concentrația cationilor divalenți Mg2+ și Ca2+ este reglată și în celulă, tot cu consumul de ATP.
Astfel, transportul activ al glucozei, care pătrunde simultan în celulă împreună cu fluxul ionului de Na+ transportat pasiv, va depinde de activitatea pompei (K+ + Na+). Dacă această pompă (K+-Na+)- este blocată, atunci diferența de concentrație de Na+ pe ambele părți ale membranei va dispărea, difuzia Na+ în celulă va scădea și, în același timp, fluxul de glucoză în celulă. se va opri. De îndată ce activitatea (K+-Na+)-ATPazei este restabilită și se creează o diferență în concentrația ionilor, fluxul difuz de Na+ crește imediat și, în același timp, transportul glucozei. În mod similar, fluxul de aminoacizi are loc prin membrană, care sunt transportați de proteine transportoare speciale care funcționează ca sisteme de transport, transportând simultan ioni.
Transportul activ al zaharurilor și aminoacizilor în celulele bacteriene este cauzat de un gradient de ioni de hidrogen. Însăși participarea proteinelor membranare speciale implicate în transportul pasiv sau activ al compușilor cu greutate moleculară mică indică specificitatea ridicată a acestui proces. Chiar și în cazul transportului pasiv de ioni, proteinele „recunosc” un anumit ion, interacționează cu acesta și se leagă
în mod specific, își schimbă conformația și funcția. În consecință, chiar și în exemplul transportului de substanțe simple, membranele acționează ca analizoare și receptori. Acest rol de receptor este evident mai ales atunci când celula absoarbe biopolimeri.
Introducere
Transportul pe membrană este transportul de substanțe prin membrana celulară în interiorul sau în afara celulei, realizat folosind diverse mecanisme - difuzie simplă, difuzie facilitată și transport activ.
Cea mai importantă proprietate a unei membrane biologice este capacitatea sa de a trece diferite substanțe în și din celulă. Acest lucru este de mare importanță pentru autoreglare și menținerea unei compoziții celulare constante. Această funcție a membranei celulare este îndeplinită datorită permeabilității selective, adică capacității de a lăsa unele substanțe să treacă și nu altele.
Transport pasiv
Există transport pasiv și activ. Transportul pasiv are loc fără consum de energie de-a lungul unui gradient electrochimic. Cele pasive includ difuzia (simpla si facilitata), osmoza, filtrarea. Transportul activ necesită energie și are loc împotriva concentrației sau a gradienților electrici.
Tipuri de transport pasiv
Tipuri de transport pasiv de substanțe:
- Difuziune simplă
- · Osmoza
- Difuzia ionică
- Difuzare facilitată
Difuziune simplă
Difuzia este procesul prin care gazul sau substanțele dizolvate se răspândesc pentru a umple întregul volum disponibil.
Moleculele și ionii dizolvați într-un lichid se află în mișcare haotică, ciocnind între ele, moleculele de solvent și membrana celulară. Ciocnirea unei molecule sau a unui ion cu o membrană poate avea două rezultate: molecula fie va „sări” de pe membrană, fie va trece prin ea. Când probabilitatea acestui din urmă eveniment este mare, se spune că membrana este permeabilă la substanță.
Dacă concentrația unei substanțe pe ambele părți ale membranei este diferită, are loc un flux de particule, direcționat dintr-o soluție mai concentrată către una diluată. Difuzia are loc până când concentrația substanței pe ambele părți ale membranei este egalată. Atât substanțele care sunt foarte solubile în apă (hidrofile), cât și substanțele hidrofobe care sunt slab sau complet insolubile în ea trec prin membrana celulară.
Substanțe hidrofobe, foarte liposolubile, difuzează datorită dizolvării în lipidele membranare. Apa și substanțele care sunt foarte solubile în ea pătrund prin defecte temporare în regiunea hidrocarburică a membranei, așa-numita. îndoituri, precum și prin pori, zonele hidrofile existente permanent ale membranei.
Când membrana celulară este impermeabilă sau slab permeabilă la solut, dar permeabilă la apă, este supusă forțelor osmotice. La o concentrație mai mică a unei substanțe în celulă decât în mediu, celula se micșorează; dacă concentrația de substanțe dizolvate în celulă este mai mare, apa intră în grabă în celulă.
Osmoza este mișcarea moleculelor de apă (solvent) printr-o membrană dintr-o zonă inferioară într-o zonă de concentrație mai mare a unei substanțe dizolvate. Presiunea osmotică este presiunea minimă care trebuie aplicată unei soluții pentru a preveni curgerea solventului prin membrană într-o soluție cu o concentrație mai mare a substanței.
Moleculele de solvent, ca și moleculele oricărei alte substanțe, sunt puse în mișcare de o forță care apare ca urmare a unei diferențe de potențiale chimice. Când o substanță se dizolvă, potențialul chimic al solventului scade. Prin urmare, într-o zonă în care concentrația de solut este mai mare, potențialul chimic al solventului este mai scăzut. Astfel, moleculele de solvent, trecând de la o soluție cu o concentrație mai mică la o soluție cu o concentrație mai mare, se deplasează în sens termodinamic „în jos”, „de-a lungul gradientului”.
Volumul celulelor este în mare măsură reglat de cantitatea de apă pe care o conțin. Celula nu este niciodată în echilibru complet cu mediul său. Mișcarea continuă a moleculelor și ionilor prin membrana plasmatică modifică concentrația de substanțe din celulă și, în consecință, presiunea osmotică a conținutului acesteia. Dacă o celulă secretă o substanță, atunci pentru a menține o presiune osmotică constantă trebuie fie să secrete o cantitate adecvată de apă, fie să absoarbă o cantitate echivalentă dintr-o altă substanță. Deoarece mediul care înconjoară majoritatea celulelor este hipotonic, este important ca celulele să împiedice pătrunderea unor cantități mari de apă în ele. Menținerea unui volum constant chiar și într-un mediu izotonic necesită un consum de energie, prin urmare în celulă concentrația de substanțe incapabile de difuziune (proteine, acizi nucleici etc.) este mai mare decât în mediul pericelular. În plus, metaboliții se acumulează constant în celulă, ceea ce perturbă echilibrul osmotic. Necesitatea de a cheltui energie pentru a menține un volum constant este ușor de demonstrat în experimente cu inhibitori de răcire sau metabolici. În astfel de condiții, celulele se umflă rapid.
Pentru a rezolva „problema osmotică”, celulele folosesc două metode: pompează componentele conținutului lor sau apa care le intră în interstițiu. În cele mai multe cazuri, celulele folosesc prima opțiune - pomparea substanțelor, cel mai adesea ioni, folosind o pompă de sodiu (vezi mai jos).
În general, volumul celulelor care nu au pereți rigizi este determinat de trei factori:
- a) cantitatea de substanțe conținute în ele care nu pot pătrunde în membrană;
- b) concentraţia în interstiţiu a compuşilor capabili să treacă prin membrană;
- c) raportul dintre ratele de penetrare și pompare a substanțelor din celulă.
Un rol important în reglarea echilibrului de apă dintre celulă și mediu îl joacă elasticitatea membranei plasmatice, care creează presiune hidrostatică care împiedică curgerea apei în celulă. Dacă există o diferență de presiune hidrostatică în două zone ale mediului, apa poate fi filtrată prin porii barierei care separă aceste zone.
Fenomenele de filtrare stau la baza multor procese fiziologice, cum ar fi formarea urinei primare în nefron, schimbul de apă între sânge și lichidul tisular din capilare.
Difuzia ionică
Difuzia ionilor are loc în principal prin structuri proteice membranare specializate - canale ionice, atunci când acestea sunt în stare deschisă. În funcție de tipul de țesut, celulele pot avea un set diferit de canale ionice. Există canale de sodiu, potasiu, calciu, sodiu-calciu și clorură. Transportul ionilor prin canale are o serie de caracteristici care îl deosebesc de difuzia simplă. Acest lucru se aplică în cea mai mare măsură canalelor de calciu.
Canalele ionice pot fi în stări deschise, închise și inactivate. Trecerea unui canal de la o stare la alta este controlată fie de o modificare a diferenței de potențial electric de pe membrană, fie de interacțiunea substanțelor active fiziologic cu receptorii. În consecință, canalele ionice sunt împărțite în dependente de tensiune și dependente de receptor. Permeabilitatea selectivă a unui canal ionic pentru un ion specific este determinată de prezența unor filtre selective speciale la gura acestuia.
Difuzare facilitată
Pe lângă apă și ioni, multe substanțe (de la etanol la medicamente complexe) pătrund prin membranele biologice prin difuzie simplă. În același timp, chiar și moleculele polare relativ mici, de exemplu, glicolii, monozaharidele și aminoacizii, practic nu pătrund în membrana majorității celulelor din cauza difuziei simple. Transferul lor se realizează prin difuzie facilitată. Difuzarea facilitată a unei substanțe de-a lungul gradientului său de concentrație, care se realizează cu participarea unor molecule speciale de transport de proteine, se numește difuzie.
Transportul Na+, K+, Cl-, Li+, Ca2+, HCO3- și H+ poate fi, de asemenea, efectuat de către purtători specifici. Trăsăturile caracteristice ale acestui tip de transport membranar sunt rata mare de transfer de substanță în comparație cu difuzia simplă, dependența de structura moleculelor sale, saturația, competiția și sensibilitatea la inhibitori specifici - compuși care inhibă difuzia facilitată.
Toate caracteristicile enumerate ale difuziei facilitate sunt rezultatul specificității proteinelor purtătoare și al numărului lor limitat în membrană. Când se atinge o anumită concentrație a substanței transportate, când toți purtătorii sunt ocupați de molecule sau ioni transportați, creșterea sa în continuare nu va duce la creșterea numărului de particule transportate - un fenomen de saturație. Substanțele care sunt similare ca structură moleculară și transportate de același purtător vor concura pentru purtător - un fenomen de competiție.
Există mai multe tipuri de transport de substanțe prin difuzie facilitată
Uniport, atunci când moleculele sau ionii sunt transportați prin membrană indiferent de prezența sau transferul altor compuși (transportul glucozei, aminoacizilor prin membrana bazală a celulelor epiteliale);
Symport, în care transferul lor are loc simultan și unidirecțional cu alți compuși (transport dependent de sodiu al zaharurilor și aminoacizilor Na+ K+, 2Cl- și cotrans-sport);
Antiport -- (transportul unei substanțe se datorează transportului simultan și în direcția opusă a unui alt compus sau ion (schimburi Na+/Ca2+, Na+/H+ Cl-/HCO3-).
Simport și antiport sunt tipuri de cotransport în care viteza de transfer este controlată de toți participanții la procesul de transport.
Natura proteinelor de transport este necunoscută. Pe baza principiului lor de funcționare, acestea sunt împărțite în două tipuri. Purtătorii de primul tip efectuează mișcări de navetă prin membrană, în timp ce purtătorii de al doilea tip sunt încorporați în membrană, formând un canal. Acțiunea lor poate fi modelată folosind antibiotice ionofore care transportă metale alcaline. Deci, unul dintre ei - (valinomicina) - acționează ca un adevărat purtător care transportă potasiul prin membrană. Moleculele de gramicidin A, un alt ionofor, sunt introduse în membrană una după alta, formând un „canal” pentru ionii de sodiu.
Majoritatea celulelor au un sistem de difuzie facilitat. Cu toate acestea, lista metaboliților transportați prin acest mecanism este destul de limitată. Acestea sunt în principal zaharuri, aminoacizi și unii ioni. Compușii care sunt produse metabolice intermediare (zaharuri fosforilate, produse ale metabolismului aminoacizilor, macroergi) nu sunt transportați folosind acest sistem. Astfel, difuzia facilitată servește la transportul acelor molecule pe care celula le primește din mediu. O excepție este transportul moleculelor organice prin epiteliu, care va fi discutat separat.
Transportul pe membrană este tranziția ionilor și moleculelor unei substanțe printr-o membrană din mediu în celulă și în direcția opusă.
În funcție de natura legăturii dintre transportul unui ion sau moleculă și transferul altor ioni și molecule, se disting următoarele:
1) uniport – transport indiferent de transportul altor legături;
2) cotransport – transport coordonat (interdependent) prin membrană; include simport (transportul simultan și unidirecțional a două substanțe diferite) și antiport (transportul simultan prin membrană în direcții opuse).
În funcție de modificarea energiei libere a sistemului, se disting două tipuri de transport:
Transport pasiv (difuzie simplă).
Transport activ– transfer de non-electroliți și ioni împotriva unui gradient chimic. sau electrochimic potențialul asociat costurilor energetice (transferul de aminoacizi și monozaharide prin membrană).
31. Transport pasiv. Fick, ecuația Nernst-Planck, Teorella.
Transport pasiv– transferul non-electroliților și ionilor prin membrană de-a lungul unui gradient chimic. sau electrochimic potențial, însoțit de o scădere a energiei libere (difuzie simplă).
Forța motrice a difuziei simple este diferența de substanță chimică. potențialele unei substanțe date în două regiuni între care are loc difuzia. Chim. potențial – o valoare egală numeric cu energia liberă pe 1 mol de substanță; este definită ca derivată parțială a energiei libere.
Principiul termodinamic de bază care guvernează distribuția în stare staționară a moleculelor care difuzează într-un sistem cu o membrană este că potențialele chimice ale unei substanțe date de ambele părți ale membranei trebuie să fie egale.
Dacă dn moli ai unei substanțe sunt transferați prin compartimentele de separare a membranei I și II, atunci acest proces este însoțit de o modificare a energiei libere a sistemului cu cantitatea:
dG = (II - I) dn.
Difuzia se oprește și sistemul intră într-o stare de echilibru termodinamic când II = I.
Prima lege a lui Fick are forma:
Fluxul unei substanțe poate fi reprezentat luând în considerare coeficientul de permeabilitate (P) al membranei pentru o substanță dată:
![]() ,
,
unde c I și c II sunt concentrațiile substanței care difuzează în soluția apoasă. [P] = cm/s.
Coeficientul de permeabilitate depinde de proprietățile membranei și ale substanțelor transferate:
unde D este coeficientul de difuzie, este coeficientul de distribuție al substanței între soluția apoasă și membrană, care caracterizează solubilitatea substanței în faza lipidică a membranei, d este grosimea membranei.
Forța motrice pentru fluxul pasiv de ioni prin membrană este gradientul de potențial electrochimic. Potențial electrochimic ion pentru condițiile în care activitatea ionului corespunde concentrației sale (c) este egală cu:
unde este potențialul electric, z este valența ionului, F este numărul Faraday, 0 este potențialul chimic standard.
Potențialul electrochimic este o măsură a muncii necesare pentru a transfera 1 mol dintr-o soluție de o concentrație dată și un potențial electric dat la un punct la infinit în vid. Această lucrare constă în costurile depășirii forțelor de interacțiune chimică (0 + RTlnc) și transferul sarcinilor în câmpul electric (zF).
Difuzia ionilor în soluție și într-o membrană neîncărcată omogenă este descrisă de ecuația de electrodifuzie Nernsta-Planck:
![]() ,
,
unde u este mobilitatea ionilor, D = uRT. Primul termen din partea dreaptă a ecuației descrie difuzia liberă, al doilea descrie migrarea ionilor într-un câmp electric.
Ecuația Teorell: Densitatea fluxului în timpul transportului pasiv: J = - cU(dm/dx), unde m este potențialul electrochimic, U este mobilitatea particulelor, c este concentrația.
32. Tipuri de transport pasiv prin membrană. Difuzare simplă și facilitată.
Transport pasiv este transferul de non-electroliți și ioni de-a lungul unei membrane de-a lungul unui gradient de potențial chimic sau electrochimic, însoțit de o scădere a energiei libere. Transportul pasiv include difuzie simplă prin bistratul lipidic şi difuzie facilitată prin canale din membrană şi cu ajutorul transportorilor. Procesele de difuzie simplă și facilitată au ca scop egalizarea gradienților și stabilirea echilibrului în sistem.
Difuzie- deplasarea spontană a unei substanțe din locuri cu o concentrație mai mare în locuri cu o concentrație mai mică a unei substanțe din cauza mișcării termice haotice a moleculelor.
Diferențele dintre difuzia facilitată și difuzia simplă:
1) transferul unei substanțe cu participarea unui purtător are loc mai rapid;
2) difuzia facilitată are proprietatea de saturație: odată cu creșterea concentrației pe o parte a membranei, densitatea de flux a substanței crește doar până la o anumită limită, când toate moleculele purtătoare sunt deja ocupate;
3) cu difuzie facilitată se observă competiţia între substanţele transportate în cazurile în care transportatorul transportă diferite substanţe; Cu toate acestea, unele substanțe sunt mai bine tolerate decât altele, iar adăugarea unor substanțe complică transportul altora; Astfel, printre zaharuri, glucoza este mai bine tolerată decât fructoza, fructoza este mai bună decât xiloza;
33. Canale ionice: mecanism de funcționare, selectivitate.
Canalele ionice sunt glicoproteine integrale care, ca urmare a influențelor externe (modificări ale potențialului membranei), pot modifica permeabilitatea membranei la diverși ioni. Canalele ionice asigură implementarea celor mai importante procese fiziologice: transmiterea semnalelor electrice și chimice, contracție, secreție.
Canalele ionice ale biomembranelor se caracterizează prin permeabilitatea selectivă pentru ioni (selectivitate) și capacitatea de a se deschide și închide sub diferite influențe asupra membranei. - Mecanismul de „gating” al canalelor este controlat de un senzor extern de stimul (receptor mesager primar).
Canalele ionice operează prin mecanismul difuziei facilitate. Mișcarea ionilor prin ei atunci când canalele sunt activate urmează un gradient de concentrație. Viteza de mișcare prin membrană este de 10 ioni pe secundă. Selectivitatea canalului este determinată de prezența unui filtru selectiv. Rolul său este jucat de secțiunea inițială a canalului, care are o anumită sarcină, configurație și dimensiune (diametru), care permite doar un anumit tip de ioni să treacă în canal. Unele dintre canalele ionice sunt neselective, cum ar fi canalele de scurgere. Acestea sunt canale membranare prin care ionii K+ părăsesc celula în stare de repaus de-a lungul unui gradient de concentrație, dar prin aceste canale o cantitate mică de ioni de Na+ intră și în celulă în stare de repaus de-a lungul unui gradient de concentrație.
34. Familii majore de canale ionice.
Un canal ionic este o proteină integrală care formează un por în membrană pentru schimbul de ioni K + , Na + , H + , Ca 2+ , Cl - , precum și apa cu mediul și este capabil să-și schimbe permeabilitatea. .
Canalele de sodiu au o structură simplă: o proteină din trei subunități diferite care formează o structură asemănătoare unui por - adică un tub cu lumen intern. Canalul poate fi în trei stări: închis, deschis și inactivat (închis și inexcitabil). Acest lucru este asigurat de localizarea sarcinilor negative și pozitive în proteina însăși; aceste sarcini sunt atrase de cele opuse existente pe membrană și astfel canalul se deschide și se închide atunci când starea membranei se schimbă. Când este deschis, ionii de sodiu pot pătrunde liber prin ea în celulă de-a lungul unui gradient de concentrație.
Canale de potasiu au o structură mai simplă: acestea sunt subunități individuale care au o formă trapezoidală în secțiune transversală; sunt situate aproape unul de celălalt, dar există întotdeauna un decalaj între ele. Canalele de potasiu nu se închid complet; în stare de repaus, potasiul părăsește liber citoplasma (de-a lungul gradientului de concentrație).
Canale de calciu- Acestea sunt proteine transmembranare cu structură complexă, formate din mai multe subunități. Prin aceste canale intră și ionii de sodiu, bariu și hidrogen. Există canale de calciu dependente de tensiune și de receptori. Prin canalele dependente de tensiune, ionii de Ca 2+ trec prin membrană de îndată ce potențialul său scade sub un anumit nivel critic. În al doilea caz, fluxul de Ca 2+ prin membrane este reglat de agonişti specifici (acetilcolină, catecolamine, serotonină, histamina etc.) în timpul interacţiunii acestora cu receptorii celulari. În prezent, există mai multe tipuri de canale de calciu cu proprietăți diferite (conductivitate, durată de deschidere) și având o localizare tisulară diferită.
Există transfer activ și pasiv (transport) de molecule neutre și ioni prin biomembrane. Transportul activ - apare atunci când energia este cheltuită din cauza hidrolizei ATP sau a transferului de protoni de-a lungul lanțului respirator mitocondrial. Transportul pasiv nu este asociat cu consumul de energie chimică de către celulă: el are loc ca urmare a difuzării substanțelor către un potențial electrochimic mai scăzut.
Un exemplu de transport activ este transferul ionilor de potasiu și sodiu prin membranele citoplasmatice K - în celulă și Na - din aceasta, transferul de calciu prin reticulul sarcoplasmatic al mușchilor scheletici și cardiaci în veziculele reticulului, transferul de ionii de hidrogen prin membranele mitocondriilor din matrice - afară: toate aceste procese au loc datorită energiei hidrolizei ATP și sunt efectuate de enzime speciale - fazele de transport ATP. Cel mai cunoscut exemplu de transport pasiv este mișcarea ionilor și a potasiului prin membrana citoplasmatică a fibrelor nervoase în timpul propagării unui potențial de acțiune.
Transferul pasiv de substanțe prin biomembrane.Difuzia moleculelor neîncărcate.
Se obișnuiește să se distingă următoarele tipuri de transfer pasiv de substanțe (inclusiv ioni) prin membrane:
2. Transfer prin pori (canale)
3. Transport folosind vectori datorită:
a) difuzia purtătorului împreună cu substanța în membrană (purtător mobil);
b) transferul de releu al unei substanțe de la o moleculă purtătoare la alta, moleculele purtătoare formează un lanț temporar peste membrană.
Transportul prin mecanismele 2 și 3 se numește uneori difuzie facilitată.
Transportul neelectroliților prin simplă șidifuzie facilitată
Diverse substanțe sunt transportate prin membrane prin două mecanisme principale: prin difuzie (transport pasiv) și prin transport activ. Permeabilitatea membranelor la diferite substanțe dizolvate depinde de mărimea și încărcătura acestor molecule. Deoarece regiunea interioară a membranei este compusă din lanțuri de hidrocarburi, multe molecule mici neutre și nepolare pot trece prin membrana bimoleculară prin difuzie obișnuită. În caz contrar, putem spune că aceste molecule sunt solubile în membrană.
Cea mai importantă dintre aceste substanțe este glucoza, care este transportată prin membrane numai în complex cu o moleculă purtătoare. Proteinele joacă de obicei acest rol. Complexul glucoză-transportor este ușor solubil în membrană și, prin urmare, poate difuza prin membrană. Acest proces se numește difuzie facilitată . Rata totală de transport al glucozei crește brusc în prezența hormonului insulină. Nu este încă pe deplin clar dacă efectul insulinei este de a crește concentrația transportorului sau dacă acest hormon stimulează formarea unui complex între glucoză și transportor.
Principalul mecanism de transport pasiv al substanțelor, datorită prezenței unui gradient de concentrație, este difuzia.
Difuzie este un proces spontan de pătrundere a unei substanțe dintr-o zonă de mai mare într-o zonă de concentrație mai mică, ca urmare a mișcării haotice termice a moleculelor.
Descrierea matematică a procesului de difuzie de către Rick. Conform legii lui Rick, viteza de difuzie este direct proporțională cu gradientul de concentrație și aria S, prin care are loc difuzia:
Semnul minus din partea dreaptă a ecuației indică faptul că difuzia are loc dintr-o zonă de concentrație mai mare într-o zonă de concentrație mai mică a substanței.
"D" numit coeficientul de difuzie . Coeficientul de difuzie este numeric egal cu cantitatea de substanță care se difuzează pe unitatea de timp printr-o unitate de suprafață cu un gradient de concentrație egal cu unitatea. „D” depinde de natura substanței și de temperatură. Caracterizează capacitatea unei substanțe de a difuza.
Deoarece gradientul de concentrație al unei membrane celulare este dificil de determinat, pentru a descrie difuzia substanțelor prin membranele celulare, se utilizează o ecuație mai simplă propusă de Kolleider și Berlund:
Unde C 1Și C 2- concentrații ale substanței pe diferite părți ale membranei, R- coeficient de permeabilitate, similar coeficientului de difuzie. Spre deosebire de coeficientul de difuzie, care depinde numai de natura substanței și de temperatură, "R" depinde și de proprietățile membranei și de starea ei funcțională.
Pătrunderea particulelor dizolvate cu sarcină electrică prin membrana celulară depinde nu numai de gradientul de concentrație al membranei. În acest sens, transportul ionilor poate avea loc în direcția opusă gradientului de concentrație, în prezența unui gradient electric direcționat opus. Combinația de concentrație și gradienți electrici se numește gradient electrochimic. Transportul pasiv al ionilor prin membrane are loc întotdeauna de-a lungul unui gradient electrochimic.
Principalii gradienți inerenți organismelor vii sunt gradienții de presiune a fluidelor de concentrație, osmotice, electrice și hidrostatice.
În conformitate cu acest gradient, există următoarele tipuri de transport pasiv de substanțe în celule și țesuturi: difuzie, osmoză, electroosmoză și osmoză anormală, filtrare.
De mare importanță pentru viața celulelor este fenomenul de transport conjugat de substanțe și ioni, care constă în faptul că transferul unei substanțe (ion) împotriva potențialului electrochimic („în sus”) este cauzat de transferul simultan al alteia. ion prin membrană în direcția scăderii potențialului electrochimic (“în jos”)”. Acest lucru este prezentat schematic în figură. Lucrarea de transport ATPazelor și transferul de protoni în timpul funcționării lanțului respirator al mitocondriilor este adesea numit transport activ primar, iar transferul de substanțe asociate cu acesta se numește transport activ secundar.
Fenomenul transferului. Ecuația generală de transport.
Un grup de fenomene cauzate de mișcarea haotică a moleculelor și care conduc la transferul de masă, energie cinetică și impuls se numește fenomen de transfer .
Acestea includ difuzia - transferul de materie, conductivitatea termică - transferul de energie cinetică și frecarea internă - transferul de impuls.
Ecuația generală de transport care descrie aceste fenomene poate fi obținută pe baza teoriei cinetice moleculare.
Lasă o anumită cantitate fizică să fie transferată printr-o zonă a zonei „S” (figura) ca rezultat al mișcării haotice a moleculelor.
La distanțe egale cu calea liberă medie, la dreapta și la stânga șantierului vom construi paralelipipedi dreptunghiulare de grosime mică " l» ( l<< ). Объем каждого параллелепипеда равен
V = Sl.
Dacă concentrația moleculelor este " P", apoi în interiorul paralelipipedului selectat există " S l p» molecule.
Toate moleculele, datorită mișcării lor haotice, pot fi reprezentate în mod convențional în șase grupuri, fiecare dintre acestea mișcându-se de-a lungul sau împotriva direcției uneia dintre axele de coordonate. Adică în direcția perpendiculară pe site " S„, moleculele se mișcă. Deoarece volumul „1” este situat la o distanță de site „ S„, atunci aceste molecule vor ajunge la el fără a se ciocni. Același număr de molecule vor ajunge la locul " S"la stânga.
Fiecare moleculă este capabilă să transfere o anumită valoare „Z” (masă, impuls, energie cinetică) și toate moleculele - sau, unde H=nZ- o mărime fizică purtată de molecule cuprinse într-o unitate de volum. Ca urmare, prin platforma " S» din volumele 1 și 2 pe perioada de timp „Dt” se transferă valoarea
Pentru a determina timpul „Dt”, presupunem că toate moleculele din volumele selectate se mișcă cu aceleași viteze medii. Apoi moleculele din volumul 1 sau 2, ajungând la locul " S", traversează-l pe o perioadă de timp
Împărțind (1) la (2), obținem că valoarea transferată în intervalul de timp „Dt” este egală cu
Modificarea valorii „H” pe unitatea de lungime „dx” se numește gradientul valorii „H”. Deoarece (H 1 - H 2) este modificarea în „H” la o distanță egală cu 2, atunci
După înlocuirea (4) în (3) și înmulțirea ecuației rezultate cu timp, găsim curgerea mărimii fizice intolerabile „H” în perioada de timp „Dt” prin zona „S”:
Aceasta este o ecuație generală de transport utilizată în studiul difuziei, conductivității termice și vâscozității.
Difuzia. Transferul pasiv al non-electroliților prin biomembrane,Ecuația lui Rick. Transportul non-electroliților prin membrane prindifuzie simplă și facilitată (în combinație cu un purtător)..
Difuzia este un proces care duce la o scădere spontană a gradienților de concentrație într-o soluție până când se stabilește o distribuție uniformă a particulelor. Procesul de difuzie joacă un rol important în multe sisteme chimice și biologice. Difuzia, de exemplu, determină în principal accesul dioxidului de carbon la structurile fotosintetice active din cloroplaste. Pentru a înțelege transportul moleculelor dizolvate prin membranele celulare, sunt necesare informații detaliate despre difuzie. Să ne uităm la câteva principii de bază ale difuziei în soluții.
Să ne imaginăm un vas, pe partea stângă a căruia se află un solvent pur, iar pe partea dreaptă există o soluție preparată cu același solvent. Mai întâi, aceste două părți ale vasului să fie separate printr-un perete vertical plat. Dacă acum scoatem peretele, atunci datorită mișcării aleatorii a moleculelor în toate direcțiile, granița dintre soluție și solvent se va deplasa spre stânga până când întregul sistem devine omogen. În 1855, Rick, în timp ce studia procesele de difuzie, a descoperit că viteza de difuzie, adică numărul de molecule de solut „n” care traversează un plan vertical pe unitatea de timp, este direct proporțională cu aria secțiunii transversale „S” și gradient de concentrație. Prin urmare,
Unde D- coeficientul de difuzie (măsurat în m 2 / s în „SI”). Semnul minus indică faptul că difuzia are loc dintr-o zonă de concentrație mare într-o zonă de concentrație scăzută. Aceasta înseamnă că gradientul de concentrație în direcția de difuzie este negativ. Ecuația (1) este cunoscută ca prima lege a difuziei a lui Rick. Legile fizice sunt concluzii intuitive care nu pot fi deduse din afirmații mai simple și ale căror consecințe nu contrazic experimentul. Astfel de concluzii includ legile mecanicii și termodinamicii; la fel și legea lui Rick.
Să luăm acum în considerare procesul de difuzie mai detaliat. Să selectăm în spațiu elementul de volum " S×dx", așa cum se arată în imagine
Viteza cu care moleculele unei substanțe dizolvate intră într-un element de volum prin secțiunea transversală „x” este egală cu Rata de modificare a gradientului de concentrație pe măsură ce „x” se modifică este egală cu
Prin urmare, viteza cu care moleculele unei substanțe dizolvate părăsesc un element de volum printr-o secțiune îndepărtată de primul cu „dx” este egală cu
Rata de acumulare a moleculelor de solut într-un element de volum este diferența dintre aceste două cantități:
Cu toate acestea, aceeași rată de acumulare a particulelor este egală cu , așa că putem scrie
Ecuația (6) numită ecuația difuziei sau a doua lege a difuziei a lui Rick, din care rezultă că modificarea concentrației în timp la o anumită distanță „x” de planul inițial este proporțională cu rata de modificare a gradientului de concentrație în direcția „x” la momentul „t”.
Pentru a rezolva ecuația (6), este necesar să folosiți metode speciale (dezvoltate de Rourier), a căror descriere este omisă; rezultatul rezultat are o formă simplă:
unde C 0 este concentrația inițială a substanței la punctul de referință la momentul zero.
Folosind ecuația (7), puteți reprezenta pe grafic dependența gradientului de concentrație de coordonatele „x” la momente diferite „t”. Prin metode optice (de exemplu, prin măsurarea indicelui de refracție) este posibil să se determine gradienții de concentrație la diferite distanțe față de limita de-a lungul căreia a început difuzia.
Mecanismul molecular al transportului ionilor activi
Există patru sisteme principale de transport ionic activ într-o celulă vie, dintre care trei asigură transferul ionilor de sodiu, potasiu, calciu și protoni prin membranele biologice datorită energiei hidrolizei ATP ca urmare a activității enzimelor purtătoare speciale numite transporta ATPaze. Cel de-al patrulea mecanism, transferul de protoni în timpul funcționării lanțului respirator mitocondrial, nu a fost încă suficient studiat. Cea mai complexă ATPază de transport este H + - ATPaza, constând din mai multe subunități, cea mai simplă este Ca 2+ ATPaza, constând dintr-un lanț polipeptidic (subunitate) cu o greutate moleculară de aproximativ 100 000. Să luăm în considerare mecanismul transportului ionilor de calciu. a acestei ATPaze.
Prima etapă a lucrării Ca 2+ ATPazei este legarea substraturilor: Ca 2+ și ATP în complex cu Mg 2+ (Mg ATP). Acești doi liganzi se atașează la locuri diferite de pe suprafața moleculei de enzimă cu fața spre exteriorul veziculului reticulului sarcoplasmatic (SR).
Ligandul este o moleculă mică (ion, hormon, medicament etc.).
A doua etapă a activității enzimei este hidroliza ATP. În acest caz, are loc formarea unui complex enzimă-fosfat (E-P).
A treia etapă a activității enzimei este tranziția centrului de legare a Ca 2+ pe cealaltă parte a membranei - translocarea.
Eliberarea energiei legăturii de înaltă energie are loc în a patra etapă a lucrului Ca 2+ ATPazei în timpul hidrolizei E-P. Această energie nu este deloc risipită (adică nu se transformă în căldură), ci este folosită pentru a schimba constanta de legare a ionilor de calciu la enzimă. Transferul de calciu de pe o parte a membranei pe cealaltă este astfel asociat cu cheltuielile de energie, care se pot ridica la 37,4 - 17,8 = 19,6 kJ/mol. Este clar că energia hidrolizei ATP este suficientă pentru a transporta doi ioni de calciu.
Transferul de calciu dintr-o zonă cu concentrații mai mici (1-4 x 10 -3 M) într-o zonă cu concentrații mai mari (1-10 x 10 -3 M) este munca prin care Ca, ATPaza de transport, face în celulele musculare.
Pentru a repeta ciclul, centrii de legare a calciului trebuie să se întoarcă din interior spre exterior, adică o altă modificare conformațională a moleculei de enzimă.
Mecanismul molecular de funcționare al acestor două „pompe” este în mare măsură similar.
Principalele etape ale Na + K + ATPaze sunt următoarele:
1. Atașarea din exterior a doi ioni K + și a unei molecule de Mg 2+ ATP:
2 K + + Mg ATP + E® (2 K +)(Mg ATP)E
2. Hidroliza ATP și formarea enzimei fosfat:
(2 K +)(Mg ATP)E® Mg ATP + (2 K +)E - P
3. Transferul centrelor de legare K+ spre interior (translocarea 1):
(2 K +)E - P ® E - P(2 K +)
4. Deconectarea ambilor ioni de potasiu și înlocuirea acestor ioni cu trei ioni de Na localizați în interiorul celulei:
E - P(2 K +) + 3 Na i + ® E - P(3 Na +) + 2 K + i
5. Hidroliza E - P:
E - P(3 Na +) ® E(3 Na +) + P (fosfat)
6. Transferul centrelor de legare împreună cu ionii Na + spre exterior (translocarea 2):
E(3Na+)® (3Na+)E
7. Eliminarea a 3 Na + și adăugarea a 2 K + din exterior:
2K0 + + 3 Na + (E)® 3 Na + + (2 K +)E
Transferul a 2 K + în celulă și eliberarea a 3 Na + în exterior duce în cele din urmă la transferul unui ion pozitiv din citoplasmă în mediu, iar acest lucru contribuie la apariția unui potențial de membrană (cu semnul minus în interiorul celulă).
Astfel, pompa Na + K + este electrogenă.
Permeabilitate
Permeabilitatea este capacitatea celulelor și țesuturilor de a absorbi, secreta și transporta substanțe chimice, trecându-le prin membranele celulare, pereții vaselor și celulele epiteliale. Celulele și țesuturile vii sunt într-o stare de schimb continuu de substanțe chimice cu mediul înconjurător, primind hrana din acesta și excretând produse metabolice în el. Principala barieră de difuzie în mișcarea substanțelor este membrana celulară. În 1899, Overton a descoperit că trecerea ușoară a substanțelor prin membrana celulară depindea de capacitatea acestor substanțe de a se dizolva în grăsimi. În același timp, o serie de substanțe polare au pătruns în celule indiferent de solubilitatea lor în grăsimi, ceea ce ar putea fi explicat prin existența porilor de apă în membrane.
În prezent, se face distincția între permeabilitatea pasivă, transportul activ al substanțelor și cazurile speciale de permeabilitate asociate cu fagocitoză și pinocitoză.
Principalele tipuri de difuzie sunt difuzia substantelor prin dizolvarea in lipidele membranare, difuzia substantelor prin porii polari si difuzia ionilor prin porii neincarcati. Sunt facilitate tipuri speciale de difuzie și schimb. Este furnizat de substanțe purtătoare speciale solubile în grăsimi care sunt capabile să lege substanța transportată pe o parte a membranei, să difuzeze cu ea prin membrană și să o elibereze pe cealaltă parte a membranei. Rolul purtătorilor de ioni specifici este îndeplinit de unele antibiotice, numite ionofori (valinomină, nigericină, monensin, antibiotice poene nistatina, aifotericina B și o serie de altele).
Ionoforii pot fi împărțiți, la rândul lor, în trei clase, în funcție de sarcina purtătorului și de structura inelului: un purtător neutru cu un inel închis de o legătură covalentă (valinomicină, nactine, poliesteri), un purtător încărcat cu un inel închis de o legătură de hidrogen (nigericină, monensin). Purtătorii încărcați au dificultăți în pătrunderea modelului și a membranelor biologice într-o formă încărcată, în timp ce într-o formă neutră difuzează liber în membrană. Forma neutră se formează prin formarea unui complex al formei anionice a purtătorului cu cationul. Astfel, purtătorii încărcați sunt capabili să schimbe cationii localizați predominant pe o parte a membranei cu cationi ai soluției de spălare pe partea opusă a membranei.
Cel mai comun tip de difuzie pasivă a membranelor celulare este porno. Datele despre proprietățile osmotice ale celulelor susțin un mecanism de permeabilitate a porilor cu adevărat existent.
Ecuația clasică a presiunii osmotice:
unde p este presiunea osmotică, c este concentrația substanței dizolvate, R este constanta gazului, T este temperatura absolută, include un termen suplimentar s, variind de la zero la 1. Această constantă, numită coeficient de reflexie, corespunde ușurința trecerii substanței dizolvate prin membrană în comparație cu trecerea unei molecule de apă.
Tipul de permeabilitate caracteristic doar celulelor și țesuturilor vii se numește transport activ. Transportul activ este transferul unei substanțe prin membrana celulară din soluția înconjurătoare (transport activ homocelular) sau prin transport activ celular, curgând împotriva gradientului activității electrochimice a substanței cu cheltuirea energiei libere a corpului. S-a dovedit acum că sistemul molecular responsabil de transportul activ al substanțelor este localizat în membrana celulară.
S-a dovedit acum că elementul principal al pompei ionice este Na + K + ATPaza. Studiul proprietăților acestei enzime membranare a arătat că enzima este doar în prezența ionilor de potasiu și sodiu, iar ionii de sodiu activează enzima din citoplasmă și ionii din soluția înconjurătoare. Un inhibitor specific al enzimei este glicozidul acid suabaina. In membranele mitocondriale se cunoaste un alt sistem molecular care asigura pomparea ionilor de hidrogen de catre enzima H + - ATPaza.
P. Mitchell, autorul teoriei chemiosmotice a fosfolării oxidative în mitocondrii, a introdus conceptul de transport activ secundar al substanțelor. Există trei metode cunoscute pentru transportul transmembranar al ionilor în membranele de împerechere. Transport unidirecțional al ionilor în direcția gradientului electrochimic prin difuzie liberă sau folosind un purtător specific - uniport. În acest din urmă caz, uniport este identic cu difuzia facilitată. O situație mai complexă apare atunci când două substanțe interacționează cu același transportor. Acest caz de simport presupune conjugarea obligatorie a fluxurilor a două substanțe în procesul de transfer al acestora prin membrană într-o singură direcție. Simportul a doi ioni este neutru din punct de vedere electric, dar echilibrul osmotic este perturbat.
Trebuie subliniat faptul că în timpul simportului, gradientul electrochimic care determină mișcarea unuia dintre ioni (de exemplu, un ion de sodiu sau un ion de hidrogen) poate provoca mișcarea unei alte substanțe (de exemplu, molecule de sazar sau aminoacizi), care este transportat de un transportator comun. Al treilea tip de conjugare ionică, actiport, caracterizează o situație în care doi ioni de același semn sunt echilibrați de-a lungul membranei în așa fel încât transferul unuia dintre ei necesită transferul celuilalt în direcția opusă. Transferul este în general neutru din punct de vedere electric și echilibrat osmotic. Acest tip de transfer este identic cu difuziunea schimbului.
Mai puțin studiate sunt două tipuri speciale de permeabilitate - fagocitoza - procesul de captare și absorbție a particulelor solide mari și pinocitoza - procesul de captare și absorbție de către o parte a suprafeței celulare a lichidului înconjurător cu substanțe dizolvate în acesta.
Toate tipurile de permeabilitate sunt, într-un grad sau altul, caracteristice țesuturilor multicelulare ale membranelor pereților vaselor de sânge, epiteliului renal, mucoasei intestinale și gastrice.
Pentru studiul permeabilității pasive și active sunt utilizate diferite metode cinetice. Cea mai utilizată metodă este metoda atomilor marcați.
Coloranții vitali sunt utilizați pe scară largă în studiile de permeabilitate. Esența metodei este de a observa, cu ajutorul unui microscop, viteza de penetrare a moleculelor de colorant în celulă. În prezent, etichetele fluorescente sunt utilizate pe scară largă, printre care fluoresceina de sodiu, clortetraciclina etc. Mult credit pentru dezvoltarea metodei de colorare vitală îi revine D.N. Nasonov, V.Ya. Alexandrov și A.S. Troshin.
Proprietățile osmotice ale celulelor și particulelor subcelulare fac posibilă utilizarea acestei calități pentru a studia permeabilitatea apei și a substanțelor solubile în ea. Esența metodei osmotice este aceea că, folosind un microscop sau măsurând împrăștierea luminii a unei suspensii de particule, se observă o modificare a volumului particulelor în funcție de tonicitatea soluției înconjurătoare.
Metodele potențiometrice sunt din ce în ce mai folosite pentru a studia membranele celulare. O gamă largă de electrozi specifici ionilor vă permite să studiați cinetica de transport a multor ioni - K +, Na +, Ca 2+, H +, CI - etc., precum și a ionilor organici - acetat, salicilați etc. .
În timpul transferului pasiv, apa, ionii și unii compuși cu molecule scăzute se mișcă liber datorită diferențelor de concentrație și egalizează concentrația substanței în interiorul și în afara celulei. În transportul pasiv, rolul principal îl joacă procesele fizice precum difuzia, osmoza și filtrarea (Fig. 24-26).
Dacă o substanță trece printr-o membrană dintr-o zonă de concentrație mare la o concentrație scăzută fără ca celula să consume energie, atunci un astfel de transport se numește pasiv sau difuziune ). Există două tipuri de difuzie: simplu Și ușoară . Membrana celulară este permeabilă la unele substanțe și impermeabilă la altele. Dacă membrana celulară este permeabilă la moleculele dizolvate, aceasta nu împiedică difuzia.
Difuziune simplă caracteristice moleculelor neutre mici (H 2 O, CO 2, O 2), precum și substanțelor organice hidrofobe cu greutate moleculară mică. Aceste molecule pot trece fără nicio interacțiune cu proteinele membranei prin porii sau canalele membranei atâta timp cât se menține gradientul de concentrație.
Difuzare facilitată. Caracteristică moleculelor hidrofile care sunt transportate prin membrană și de-a lungul unui gradient de concentrație, dar cu ajutorul unor proteine membranare speciale - purtători. Difuzia facilitată, spre deosebire de difuzia simplă, se caracterizează printr-o selectivitate ridicată, deoarece proteina transportor are un centru de legare complementar substanței transportate, iar transferul este însoțit de modificări conformaționale ale proteinei.
Unul dintre posibilele mecanisme de difuzie facilitată poate fi următorul: o proteină de transport (translocaza) leagă o substanță, apoi se apropie de partea opusă a membranei, eliberează această substanță, își ia conformația inițială și este din nou gata să îndeplinească funcția de transport. . Se știe puțin despre cum se mișcă proteina în sine. Un alt mecanism posibil de transport implică participarea mai multor proteine transportoare. În acest caz, compusul legat inițial în sine se deplasează de la o proteină la alta, legându-se secvenţial cu una sau cealaltă proteină până când ajunge pe partea opusă a membranei.
În ceea ce privește transportul ionilor, acesta se realizează, de regulă, folosind difuzie prin special canale ionice (Fig.27).
Fig.27. Principalele mecanisme de transmitere transmembranară a informaţiei semnal: I - trecerea unei molecule semnal liposolubile prin membrana celulară; II - legarea unei molecule semnal de receptor și activarea fragmentului său intracelular; III - reglarea activității canalelor ionice; IV - transmiterea informațiilor de semnal folosind emițătoare secundare. 1 - medicament; 2 - receptor intracelular; 3 - receptor celular (transmembranar); 4 - transformare intracelulară (reacție biochimică); 5 - canal ionic; 6 - flux ionic; 7 - intermediar secundar; 8 - enzimă sau canal ionic; 9 - intermediar secundar.
Astfel, există mai multe mecanisme de transport al substanțelor.
Primul mecanism este că o moleculă de semnalizare solubilă în lipide trece prin membrana celulară și activează un receptor intracelular (de exemplu, o enzimă). Acest lucru se realizează prin oxid nitric, o serie de hormoni liposolubili (glucocorticoizi, mineralocorticoizi, hormoni sexuali și hormoni tiroidieni) și vitamina D. Aceștia stimulează transcrierea genelor în nucleul celular și, astfel, sinteza de noi proteine. Mecanismul de acțiune al hormonilor este stimularea sintezei de noi proteine în nucleul celular, care rămân active în celulă pentru o perioadă lungă de timp.
Al doilea mecanism de transmitere a semnalului prin membrana celulară este legarea de receptorii celulari care au fragmente extracelulare și intracelulare (adică receptori transmembranari). Astfel de receptori sunt intermediari în prima etapă a acțiunii insulinei și a unui număr de alți hormoni. Părțile extracelulare și intracelulare ale unor astfel de receptori sunt conectate printr-o punte polipeptidică care trece prin membrana celulară. Fragmentul intracelular are activitate enzimatică, care crește atunci când molecula de semnalizare se leagă de receptor. În consecință, rata reacțiilor intracelulare la care participă acest fragment crește.
Al treilea mecanism de transfer de informații este efectul asupra receptorilor care reglează deschiderea sau închiderea canalelor ionice. Moleculele de semnalizare naturale care interacționează cu astfel de receptori includ, în special, acetilcolina, acidul gama-aminobutiric (GABA), glicina, aspartatul, glutamatul și altele, care sunt mediatori ai diferitelor procese fiziologice. Atunci când interacționează cu receptorul, conductanța transmembranară pentru ionii individuali crește, ceea ce determină o modificare a potențialului electric al membranei celulare. De exemplu, acetilcolina, interacționând cu receptorii H-colinergici, crește intrarea ionilor de sodiu în celulă și provoacă depolarizare și contracție musculară. Interacțiunea acidului gamma-aminobutiric cu receptorul său duce la creșterea aportului de ioni de clor în celule, polarizare crescută și dezvoltarea inhibării (opresiunii) a sistemului nervos central. Acest mecanism de semnalizare se caracterizează prin dezvoltarea rapidă a efectului (milisecunde).
Al patrulea mecanism de transmitere transmembranară a unui semnal chimic este realizat prin receptori care activează un transmițător secundar intracelular. Atunci când interacționează cu astfel de receptori, procesul are loc în patru etape. Molecula de semnalizare este recunoscută de un receptor de pe suprafața membranei celulare; ca urmare a interacțiunii lor, receptorul activează proteina G de pe suprafața interioară a membranei. O proteină G activată modifică activitatea fie a unei enzime, fie a unui canal ionic. Aceasta duce la o modificare a concentrației intracelulare a mesagerului secundar, prin care efectele se realizează direct (se modifică procesele metabolice și energetice). Acest mecanism de transmitere a informațiilor de semnal face posibilă întărirea semnalului transmis. Deci, dacă interacțiunea unei molecule de semnalizare (de exemplu, norepinefrina) cu receptorul durează câteva milisecunde, atunci activitatea emițătorului secundar, către care receptorul transmite semnalul, persistă zeci de secunde.
Mesagerii secundari sunt substanțe care se formează în interiorul celulei și sunt componente importante ale numeroaselor reacții biochimice intracelulare. Intensitatea și rezultatele activității celulare și funcționarea întregului țesut depind în mare măsură de concentrația acestora. Cei mai cunoscuți mesageri secundi sunt adenozin monofosfat ciclic (cAMP), guanozin monofosfat ciclic (cGMP), ionii de calciu și potasiu etc.
Osmoză- un tip special de difuzie a apei printr-o membrană semi-permeabilă într-o zonă de concentrație mai mare a unei substanțe dizolvate. Ca urmare a acestei mișcări, în interiorul celulei se creează o presiune semnificativă, care se numește presiune osmotică. Această presiune poate chiar distruge celula.
De exemplu, dacă celulele roșii din sânge sunt plasate în apă curată, atunci sub influența osmozei apa va pătrunde în ele mai repede decât va părăsi. Acest mediu se numește hipotonic. Pe măsură ce apa pătrunde, celulele roșii din sânge se vor umfla și „explodează”. O altă situație este un mediu izotonic. Dacă plasați globule roșii în apă care conține 0,87% sare de masă, nu se creează presiune osmotică. Acest lucru se explică prin faptul că, cu concentrații egale de soluție în interiorul și în afara celulei, apa se mișcă în mod egal în ambele direcții. Un mediu este considerat hipertonic atunci când concentrația de substanțe dizolvate în el este mai mare decât în celulă. O celulă (eritrocit) într-un astfel de mediu începe să piardă apă, se micșorează și moare.
Toate aceste caracteristici ale osmozei sunt luate în considerare la administrarea substanțelor medicinale. De regulă, medicamentele destinate injectării sunt preparate într-o soluție izotonă. Acest lucru previne umflarea sau micșorarea celulelor sanguine atunci când medicamentul este administrat. Picăturile nazale se prepară și într-o soluție izotonă pentru a evita umflarea sau deshidratarea celulelor mucoasei nazale.
Osmoza explică și unele dintre efectele medicamentelor, de exemplu, efectul laxativ al sărurilor Epsom (sulfat de magneziu) și al altor laxative saline. În lumenul intestinal formează un mediu hipertonic. Sub influența osmozei, apa părăsește celulele epiteliale intestinale, spațiul intercelular și sângele în lumenul intestinal, întinde pereții intestinali, își lichefiază conținutul și accelerează golirea.
Filtrare- mișcarea moleculelor de apă și a substanțelor dizolvate în ea prin membrana celulară în direcția opusă acțiunii presiunii osmotice.
Acest proces devine posibil dacă soluția din celulă este sub o presiune mai mare decât osmotică. De exemplu, inima pompează sânge în vase sub o anumită presiune. În cele mai subțiri capilare, această presiune crește și devine suficientă pentru a forța apa și substanțele dizolvate în sânge să părăsească capilarele în spațiul intercelular. Se formează așa-numitul fluid tisular; acesta joacă un rol important în furnizarea de nutrienți către celule și în îndepărtarea produselor finite metabolice din acestea. După îndeplinirea funcțiilor sale, lichidul tisular sub formă de limfă revine în fluxul sanguin prin vasele limfatice.
De asemenea, filtrarea joacă un rol important în funcționarea rinichilor. În capilarele rinichilor, sângele este sub presiune mare, ceea ce determină filtrarea apei și a substanțelor dizolvate în ea din vasele de sânge în cei mai subțiri tubuli renali. Apoi o parte din apa și substanțele necesare organismului sunt din nou absorbite și intră în fluxul sanguin general, iar partea rămasă formează urină și este excretată din organism.