Membrana plasmatica: structura si functii. Membrana plasmatica Structura membranei plasmatice
Secțiuni: Biologie
Articolul este un rezumat al unei lecții-studiu și al consolidării primare a noilor cunoștințe (cursul „Biologie generală”, nota 10, conform programului lui V.B. Zakharov).
Sarcini:
- formarea cunoștințelor despre structura, proprietățile și funcțiile stratului interior al membranei celulare - membrana plasmatică (și, folosind exemplul acesteia, alte membrane celulare), folosind un balon de săpun ca model.
- dezvoltarea conceptului de corespondenţă a structurii cu funcţiile îndeplinite.
- consolidarea primară a cunoștințelor dobândite folosind sarcini în formatul Unified State Exam.
Echipament:
- tabelul „Structura celulelor vegetale și animale în funcție de microscoape luminoase și electronice”.
- soluție de detergent (pentru a produce bule de săpun), tub de plastic, ac subțire de cusut.
- desen la tablă: modele moleculare<Poza 1 >.
- materiale didactice cu sarcini în format Unified State Exam.
În timpul orelor
Profesor:În ultima lecție, am desfășurat lucrări de laborator „Plasmoliza și deplasmoliza în celulele pielii de ceapă”, în timpul cărora ne-am familiarizat cu fenomene interesante. Care este esența lor?
Elevi: Atunci când țesutul vegetal (epiderma solzilor de ceapă) a fost plasat într-o soluție hipertonă de clorură de sodiu (NaCl), nu a existat o difuzie a acestei soluții în celule, ci o eliberare de apă din vacuolele celulare către soluția hipertonică de NaCl pentru pentru a echilibra concentrațiile de ioni de pe ambele părți ale membranei celulare. În același timp, volumul vacuolelor și a întregii citoplasme a scăzut, ceea ce a dus la separarea citoplasmei de peretele celular - plasmoliza. La returnarea țesutului studiat în apă curată, nu am observat nici eliberarea de substanțe dizolvate din vacuole, ci doar curgerea apei din spațiul înconjurător în celulă, în vacuole cu seva celulară, ceea ce a dus la refacerea volumul celular la limitele sale anterioare – deplasmoliza.
Profesor: Ce concluzie se poate trage din experiment?
Elevi: Probabil, suprafața celulei permite apei să treacă în ambele direcții, dar reține ionii Na + și Cl - care fac parte din sarea de masă.
Profesor: Proprietatea pe care am descoperit-o se numește permeabilitate selectivă sau semipermeabilitatea membranei plasmatice.
Ce este o membrană plasmatică (sau membrană plasmatică), care este structura, proprietățile și funcțiile ei, trebuie să înțelegem în lecția de astăzi. După cum am convenit, lecția va fi predată de tovarășii tăi care au pregătit o prelegere despre membranele celulare. Sarcina ta este să notezi informații de bază despre membranele celulare în timp ce asculți. Va trebui să aplicați cunoștințele dobândite răspunzând la întrebarea test de la sfârșitul lecției.
Lector 1. Structura membranei.
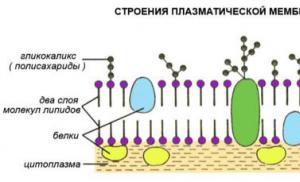
Membrana plasmatică este prezentă în toate celulele (sub glicocalix la animale și sub peretele celular la alte organisme), ea asigură interacțiunea celulei cu mediul său. Plasmalema formează o suprafață mobilă a celulei, care poate avea excrescențe și invaginări, face mișcări oscilatorii sub formă de undă, iar macromoleculele se mișcă constant în ea.
În ciuda acestor schimbări continue, celula rămâne întotdeauna înconjurată de o membrană strânsă. Membrana plasmatică este o peliculă subțire de mai puțin de 10 nm grosime. Chiar dacă grosimea sa este mărită de 1 milion de ori, vom obține o valoare de doar aproximativ 1 cm, în timp ce dacă întreaga celulă este mărită de 1 milion de ori, dimensiunea ei va fi comparabilă cu un public destul de mare.
Membrana conține două tipuri principale de molecule: fosfolipide, formând dublu stratîn grosimea membranei și veverite pe suprafetele sale. Aceste molecule sunt ținute împreună prin interacțiuni non-covalente. Acest model de membrană asemănător sandvișului a fost propus de oamenii de știință americani Danieli și Dawson în 1935. Odată cu apariția microscopului electronic, acesta a fost confirmat și oarecum modificat. Momentan acceptat model de membrană mozaic fluid, conform căreia moleculele de proteine care plutesc într-un strat dublu lipidic lichid formează un fel de mozaic în el. O diagramă a acestui model modern, propus în 1972 de Singer și Nicholson, este dată în manual.
Unele proteine au carbohidrați atașați covalent de suprafața lor exterioară, formându-se glicoproteine– antene moleculare deosebite care sunt receptori. Glicoproteinele sunt implicate în recunoașterea semnalelor externe care vin din mediul înconjurător sau din alte părți ale corpului însuși și în răspunsul celulelor la influența lor. O astfel de recunoaștere reciprocă este o etapă necesară premergătoare fertilizării, precum și aderarea celulelor în procesul de diferențiere a țesuturilor. Recunoașterea este, de asemenea, asociată cu reglarea transportului moleculelor și ionilor prin membrană, precum și cu răspunsul imun, în care glicoproteinele joacă rolul de antigene.
Lector 2.Proprietățile membranei.
Pentru a înțelege ce proprietăți au aceste structuri microscopice, să luăm ca model un balon de săpun. Faptul este că moleculele de săpun și fosfolipide care alcătuiesc membranele au o structură similară<Poza 1>. Săpunurile (sărurile acizilor grași) au în structură cap hidrofil(dintr-o grupare carboxil încărcată) și lung coadă hidrofobă. Fosfolipidele care alcătuiesc membranele au și o coadă hidrofobă (din două lanțuri de acizi grași) și un cap hidrofil mare care conține o grupă de acid fosforic încărcată negativ.
Orez. 1. Modele de molecule.
Atunci când substanțe cu o structură similară sunt amestecate cu apă, moleculele lor capătă în mod spontan următoarea configurație: capetele hidrofile sunt scufundate în apă, iar cozile hidrofobe nu intră în contact cu apa, contactându-se doar între ele și cu alte substanțe hidrofobe care poate fi în preajmă, de exemplu, cu aer. Aflându-se la granița dintre două medii de natură similară, atât moleculele de săpun, cât și moleculele de fosfolipide sunt capabile să formeze un strat dublu. Unele dintre proprietățile importante ale membranelor biologice (cum ar fi bulele de săpun), enumerate mai jos, sunt explicate prin structura stratului dublu lipidic.
A) Mobilitate.
Un strat dublu lipidic este în esență o formațiune lichidă, în planul căreia moleculele se pot mișca liber - „curge” fără a pierde contactele din cauza atracției reciproce ( „Lectorul” demonstrează fluxul de lichid în peretele unui balon de săpun atârnat pe un tub de plastic). Cozile hidrofobe pot aluneca liber una pe lângă alta.
b) Capacitate de autoblocare.
„Lectorul” demonstrează cum, atunci când un balon de săpun este străpuns și acul este îndepărtat ulterior, integritatea peretelui său este imediat restaurată. Datorită acestei abilități, celulele pot fuziona prin fuzionarea membranelor lor plasmatice (de exemplu, în timpul dezvoltării țesutului muscular). Același efect se observă atunci când tăiați o celulă în două părți cu un microcuțit, după care fiecare parte este înconjurată de o membrană plasmatică închisă.
V) Permeabilitate selectivă.
Adică impermeabilitatea la moleculele solubile în apă datorită peliculei uleioase formate de cozile hidrofobe ale moleculelor de fosfolipide. Pentru a pătrunde fizic într-un astfel de film, substanța în sine trebuie să fie hidrofobă sau se poate strecura prin goluri aleatorii formate ca urmare a mișcărilor moleculare (molecule mici, cum ar fi moleculele de apă).
Proteinele care patrund in intreaga grosime a membranei, sau sunt situate pe suprafetele sale exterioare si interioare, ajuta celulele sa faca schimb de substante cu mediul. Moleculele proteice asigură transportul selectiv al substanțelor prin membrană, fiind enzime; în plus, se formează pori în interiorul moleculelor proteice sau între moleculele vecine, prin care apa și unii ioni pătrund pasiv în celule.
Lector 3. Funcțiile membranei plasmatice.
La ce servește o structură cu o astfel de structură și proprietăți pentru o celulă? Se dovedește că ea:
- Oferă formă celulei și protejează de daune fizice și chimice.
- Datorită mobilității sale, capacității de a forma excrescențe și proeminențe, realizează contactul și interacțiunea celulelor în țesuturi și organe.
- Separă mediul celular de mediul extern și menține diferențele dintre acestea.
- Este un fel de indicator al tipului de celule datorită faptului că proteinele și carbohidrații de pe suprafața membranelor și diferitele celule nu sunt la fel.
- Reglează schimbul dintre celulă și mediu, asigurând selectiv transportul nutrienților în celulă și îndepărtarea produselor metabolice finale către exterior.
Lector 4. Vreau să vă spun cum se întâmplă transport prin membrana plasmatică, și în mod similar prin alte membrane celulare. Transportul poate fi pasiv, care nu necesită energie, și activ, dependent de energie, timp în care se consumă energie rezultată din hidroliza moleculelor de ATP.
1. Difuzia.
Acesta este un proces pasiv; mișcarea substanțelor are loc dintr-o zonă de concentrație mare într-o zonă de concentrație scăzută. Gazele și moleculele lipofile (solubile în grăsimi) difuzează rapid, ionii și moleculele polare mici (glucoză, aminoacizi, acizi grași) difuzează lent. Difuzia este accelerată de porii din moleculele de proteine.
Un tip de difuzie este osmoză– mișcarea apei prin membrană.
2. Endocitoza.
Acesta este transportul activ al substanțelor prin membrană în celulă (exocitoză - în afara celulei). În funcție de natura substanței transferate prin membrană, se disting două tipuri de aceste procese: dacă este transferată o substanță densă - fagocitoză(din grecescul „phagos” - devorare și „cytos” - celulă), dacă picături de lichid care conțin diferite substanțe în stare dizolvată sau suspendată, atunci - pinocitoza(din grecescul „pino” - băutură și „cytos” - celulă).
Principiul transferului în ambele cazuri este identic: în locul în care suprafața celulei intră în contact cu o particulă sau picătură de substanță, membrana se îndoaie, formează o depresiune și înconjoară particula sau picătura de lichid, care este scufundată. într-un „pachet cu membrană” în interiorul celulei. Aici se formează o vacuolă digestivă, iar substanțele organice care intră în celulă sunt digerate în ea. Fagocitoza este larg răspândită la animale, iar pinocitoza este efectuată de celulele animalelor, plantelor, ciupercilor, bacteriilor și algelor albastre-verzi.
3. Transport activ folosind enzime încorporate în membrană.
Transferul are loc în raport cu gradientul de concentrație cu cheltuiala de energie, de exemplu, ionii de potasiu intră („pompați”) în celulă, iar ionii de sodiu sunt îndepărtați („pompați”) din celulă. Acest lucru este însoțit de acumularea unei diferențe de potențial electric pe membrană. Astfel de sisteme de transport celular sunt de obicei numite „ pompe" Transportul aminoacizilor și zaharurilor se realizează în mod similar.
concluzii:
- Plasmalemma este o peliculă subțire, de aproximativ 10 nm grosime, pe suprafața celulei. Include structuri lipoproteice (lipide și proteine).
- Unele molecule de suprafață ale proteinelor au molecule de carbohidrați atașate (sunt asociate cu mecanismul de recunoaștere).
- Lipidele membranare formează spontan un strat dublu. Aceasta determină permeabilitatea selectivă a membranei.
- Proteinele membranei îndeplinesc o varietate de funcții și facilitează semnificativ transportul prin membrană.
- Lipidele și proteinele membranei sunt capabile să se deplaseze în planul membranei, datorită căruia suprafața celulei nu este perfect netedă.
Pentru a consolida informațiile primite în lecție, studenților li se oferă sarcini în formatul Unified State Exam.
Partea „A”
Alegeți un răspuns corect.
A1. Structura și funcțiile membranei plasmatice sunt determinate de moleculele care o alcătuiesc:
1) glicogen și amidon
2) ADN și ATP
3) proteine și lipide
4) fibre și glucoză
A2. Membrana plasmatică nu îndeplinește funcția de:
1) transportul de substante
2) protecția celulelor
3) interacțiunea cu alte celule
4) sinteza proteinelor
A3. Carbohidrații incluși în structura membranei celulare îndeplinesc următoarele funcții:
1) transportul de substante
2) receptor
3) formarea unui strat dublu de membrană
4) fotosinteza
A4. Proteinele incluse în structura membranei celulare îndeplinesc următoarele funcții:
1) construcție
2) protectoare
3) transport
4) toate funcțiile specificate
A5. Fagocitoza este:
1) absorbția lichidului de către celulă
2) captarea particulelor solide
3) transportul substanțelor prin membrană
4) accelerarea reacțiilor biochimice
A6. Suprafețele hidrofile ale membranelor sunt formate:
1) cozile nepolare ale lipidelor
2) capetele polare ale lipidelor
3) proteine
4) carbohidrați
A7. Trecerea ionilor de Na + și K + prin membrană are loc prin:
1) difuzie
2) osmoza
3) transfer activ
4) neimplementat
A8. Următoarele trec liber prin stratul lipidic al membranei:
1) apa
2) difuzat
3) glucoză
4) amidon
Partea „B”
1) transportul activ consumă energie
2) fagocitoza este un tip de endocitoză
3) difuzia este un tip de transport activ
4) peretele celular al plantelor este format din celuloză
5) osmoza este difuzia apei
6) pinocitoza este un tip de fagocitoză
7) plasmalema este formată din trei straturi de lipide
8) o celulă animală nu are perete celular
9) plasmalema asigură comunicarea între celulă și mediul ei
Partea „C”
Sarcini cu răspuns gratuit detaliat
C1. Care este semnificația endocitozei:
a) pentru protozoare și nevertebrate inferioare?
b) pentru animale și oameni foarte organizați?
C2. Care este baza fizică a transportului vacuolar într-o celulă?
C3. Care este semnificația biologică a neregulilor de pe suprafața plasmalemei unor celule (microvili, cili etc.)?
C4. Raza electrică și anghila electrică își uimesc prada cu descărcări de câteva sute de volți. Ce proprietăți ale membranelor plasmatice celulare susțin posibilitatea de a crea astfel de descărcări?
C5. Cum funcționează plasmalema pentru a oferi celulei o „carte de identitate”?
Răspunsuri la sarcini.
Partea „A”.
1–3, 2–4, 3–2, 4–4, 5–2, 6–2, 7–3, 8–2.
Partea „B”.
1, 2, 4, 5, 8, 9 – „da”; 3, 6, 7 – „nu”
Partea „C”.
1a. Posibilitatea de intrare a alimentelor în celule și digestie ulterioară în lizozomi.
1b. Activitatea fagocitară a leucocitelor este de mare importanță în protejarea organismului de bacteriile patogene și alte particule nedorite. Pinocitoza în celulele tubulare renale duce la absorbția proteinelor din urina primară.
2. Principalele proprietăți ale straturilor duble lipidice sunt capacitatea membranelor de a se închide.
3. Creșterea suprafeței celulei pentru schimbul dintre celulă și mediul său.
4. Prezența sistemelor enzimatice care efectuează transport activ („pompe”) duce la o redistribuire a sarcinilor pe plasmalemă și la crearea unei diferențe de potențial membranar.
5. Pentru aceasta, există o serie de grupuri chimice specifice pe suprafața membranei - „antene”, care sunt, cel mai adesea, glicoproteine.
Membrană biologică universală format dintr-un strat dublu de molecule de fosfolipide cu o grosime totală de 6 microni. În acest caz, cozile hidrofobe ale moleculelor de fosfolipide sunt întoarse spre interior, unele către altele, iar capetele hidrofile polare sunt întoarse spre exteriorul membranei, spre apă. Lipidele asigură proprietățile fizico-chimice de bază ale membranelor, în special ale acestora fluiditate la temperatura corpului. În acest dublu strat lipidic sunt încorporate proteine.
Ele sunt împărțite în integrală(penetrează întregul strat dublu lipidic), semi-integral(penetrează până la jumătate din stratul dublu lipidic) sau suprafața (situată pe suprafața interioară sau exterioară a stratului dublu lipidic).
În acest caz, moleculele de proteine sunt situate într-un model mozaic în stratul dublu lipidic și pot „pluti” în „marea lipidică” ca aisbergurile, datorită fluidității membranelor. În funcție de funcția lor, aceste proteine pot fi structural(menține o anumită structură a membranei), receptor(formă receptori pentru substanțele biologic active), transport(transport substante prin membrana) si enzimatic(cataliza anumite reactii chimice). Acesta este în prezent cel mai recunoscut model mozaic fluid membrana biologică a fost propusă în 1972 de Singer și Nikolson.
Membranele îndeplinesc o funcție de demarcație în celulă. Ele împart celula în compartimente, în care procesele și reacțiile chimice pot avea loc independent unele de altele. De exemplu, enzimele hidrolitice agresive ale lizozomilor, capabile să descompună majoritatea moleculelor organice, sunt separate de restul citoplasmei printr-o membrană. Dacă este distrus, are loc autodigestia și moartea celulelor.
Având un plan structural general, diferitele membrane celulare biologice diferă prin compoziția chimică, organizarea și proprietățile lor, în funcție de funcțiile structurilor pe care le formează.
Membrana plasmatica, structura, functiile.
Citolema este o membrană biologică care înconjoară celula din exterior. Aceasta este cea mai groasă (10 nm) și cea mai complexă membrană celulară organizată. Se bazează pe o membrană biologică universală acoperită la exterior glicocalixși din interior, din partea citoplasmei, stratul submembranar(Fig. 2-1B). Glicocalix(3-4 nm grosime) este reprezentată de regiunile exterioare, carbohidrate, ale proteinelor complexe - glicoproteinele și glicolipidele care alcătuiesc membrana. Aceste lanțuri de carbohidrați joacă rolul de receptori care asigură faptul că celula recunoaște celulele vecine și substanța intercelulară și interacționează cu acestea. Acest strat include, de asemenea, proteine de suprafață și semi-integrale, ale căror regiuni funcționale sunt situate în zona supramembrană (de exemplu, imunoglobuline). Glicocalixul conține receptori de histocompatibilitate, receptori pentru mulți hormoni și neurotransmițători.
Strat submembranos, cortical format din microtubuli, microfibrile și microfilamente contractile, care fac parte din citoscheletul celular. Stratul submembranar menține forma celulei, îi creează elasticitatea și asigură modificări ale suprafeței celulei. Datorită acestui fapt, celula participă la endo- și exocitoză, secreție și mișcare.
Citolema funcționează o multime de funcții:
1) delimitare (citolema separă, delimitează celula de mediu și asigură legătura acesteia cu mediul extern);
2) recunoașterea de către această celulă a altor celule și atașarea la acestea;
3) recunoașterea de către celulă a substanței intercelulare și atașarea la elementele acesteia (fibre, membrana bazală);
4) transportul de substanțe și particule în și din citoplasmă;
5) interacțiunea cu moleculele de semnalizare (hormoni, mediatori, citokine) datorită prezenței unor receptori specifici pentru acestea pe suprafața sa;
- asigură deplasarea celulară (formarea pseudopodiilor) datorită conexiunii citolemei cu elementele contractile ale citoscheletului.
Citolema conține numeroase receptori, prin care substanțe biologic active ( liganzi, molecule de semnalizare, primi mesageri: hormoni, mediatori, factori de crestere) actioneaza asupra celulei. Receptorii sunt senzori macromoleculari determinați genetic (proteine, glico- și lipoproteine) încorporați în citolemă sau localizați în interiorul celulei și specializați în perceperea unor semnale specifice de natură chimică sau fizică. Substanțele biologic active, atunci când interacționează cu un receptor, provoacă o cascadă de modificări biochimice în celulă, transformându-se într-un răspuns fiziologic specific (modificarea funcției celulare).
Toți receptorii au un plan structural general și sunt formați din trei părți: 1) supramembrană, care interacționează cu substanța (ligand); 2) intramembranar, care efectuează transferul de semnal și 3) intracelular, cufundat în citoplasmă.
Tipuri de contacte intercelulare.
Citolema este, de asemenea, implicată în formarea de structuri speciale - conexiuni intercelulare, contacte, care asigură o interacțiune strânsă între celulele adiacente. Distinge simpluȘi complex conexiuni intercelulare. ÎN simplu La joncțiunile intercelulare, citolemele celulelor se apropie de o distanță de 15-20 nm și moleculele glicocalixului lor interacționează între ele (Fig. 2-3). Uneori, proeminența citolemei unei celule intră în adâncitura unei celule adiacente, formând conexiuni zimțate și asemănătoare degetelor (conexiuni „de tip blocare”).
Complex Există mai multe tipuri de conexiuni intercelulare: blocare, interblocareȘi comunicare(Fig. 2-3). LA blocare compușii includ contact strâns sau zona de blocare. În acest caz, proteinele integrale ale glicocalixului celulelor învecinate formează un fel de rețea celulară de-a lungul perimetrului celulelor epiteliale învecinate în părțile lor apicale. Datorită acesteia, golurile intercelulare sunt închise și delimitate de mediul extern (Fig. 2-3).
Orez. 2-3. Diverse tipuri de conexiuni intercelulare.
- Conexiune simplă.
- Conexiune strânsă.
- Curea adezivă.
- Desmosom.
- Hemidesmozomul.
- Slot (comunicare) conexiune.
- Microvilozități.
(După Yu. I. Afanasyev, N. A. Yurina).
LA închegată, racordurile de ancorare includ adeziv centuraȘi desmozomi. Curea adezivă situat în jurul părților apicale ale celulelor epiteliale cu un singur strat. În această zonă, glicoproteinele integrale ale glicocalixului celulelor învecinate interacționează între ele, iar proteinele submembranare, inclusiv mănunchiuri de microfilamente de actină, se apropie de ele din citoplasmă. Desmozomi (pete de aderență)– structuri pereche cu o dimensiune de aproximativ 0,5 microni. În ele, glicoproteinele citolemei celulelor învecinate interacționează strâns, iar din partea celulelor din aceste zone, mănunchiuri de filamente intermediare ale citoscheletului celular sunt țesute în citolemă (Fig. 2-3).
LA conexiuni de comunicare include joncțiuni gap (nexus) și sinapse. Legături au o dimensiune de 0,5-3 microni. În ele, citolemele celulelor învecinate se apropie de 2-3 nm și au numeroase canale ionice. Prin intermediul acestora, ionii pot trece de la o celulă la alta, transmitând excitația, de exemplu, între celulele miocardice. Sinapsele caracteristice țesutului nervos și apar între celulele nervoase, precum și între celulele nervoase și efectoare (mușchi, glandulare). Au o despicatură sinaptică, unde, la trecerea unui impuls nervos, un neurotransmițător este eliberat din partea presinaptică a sinapsei, care transmite impulsul nervos către o altă celulă (pentru mai multe detalii, vezi capitolul „Țesut nervos”).
Prelegerea nr. 4.
Numar de ore: 2
Membrană plasmatică
1.
2.
3. Contacte intercelulare.
1. Structura membranei plasmatice
Membrana plasmatica sau plasmalema, este o structură periferică superficială care limiteazăcelulă în exterior și asigurând legătura acesteia cu alte celule și cu mediul extracelular. Are o grosimeaproximativ 10 nm. Printre alte membrane celulare, plasmalema este cea mai groasă. Din punct de vedere chimic, membrana plasmatică este complex de lipoproteine. Componentele principale sunt lipidele (aproximativ 40%), proteinele (mai mult de 60%) și carbohidrații (aproximativ 2-10%).
Lipidele includ un grup mare de substanțe organice care au o solubilitate slabă în apă (hidrofobicitate) și o solubilitate bună în solvenți organici și grăsimi (lipofilitate).Lipidele tipice găsite în membrana plasmatică sunt fosfolipidele, sfingomielinele și colesterolul. În celulele vegetale, colesterolul este înlocuit cu fitosterol. Pe baza rolului lor biologic, proteinele plasmalemei pot fi împărțite în proteine enzimatice, receptori și proteine structurale. Carbohidrații din plasmalemma fac parte din plasmalemă în stare legată (glicolipide și glicoproteine).
În prezent este general acceptat model fluid-mozaic al structurii unei membrane biologice. Conform acestui model, baza structurală a membranei este formată dintr-un strat dublu de fosfolipide încrustate cu proteine. Cozile moleculelor se confruntă într-un strat dublu, în timp ce capetele polare rămân în exterior, formând suprafețe hidrofile. Moleculele proteice nu formează un strat continuu, ele sunt situate în stratul lipidic, plonjând la diferite adâncimi (există proteine periferice, unele proteine pătrund prin membrană, altele sunt scufundate în stratul lipidic). Majoritatea proteinelor nu sunt asociate cu lipidele membranare, de exemplu. par să plutească într-un „lac lipidic”. Prin urmare, moleculele de proteine sunt capabile să se miște de-a lungul membranei, să se asambla în grupuri sau, dimpotrivă, să se împrăștie pe suprafața membranei. Acest lucru sugerează că membrana plasmatică nu este o formațiune statică, înghețată.
În afara plasmalemei există un strat supramembranar - glicocalix. Grosimea acestui strat este de aproximativ 3-4 nm. Glicocalixul se găsește în aproape toate celulele animale. Este asociat cu plasmalema complex glicoproteic. Carbohidrații formează lanțuri lungi, ramificate de polizaharide asociate cu proteinele și lipidele membranei plasmatice. Glicocalixul poate conține proteine enzimatice implicate în descompunerea extracelulară a diferitelor substanțe. Produsele cu activitate enzimatică (aminoacizi, nucleotide, acizi grași etc.) sunt transportate prin membrana plasmatică și absorbite de celule.
Membrana plasmatică este reînnoită constant. Acest lucru se întâmplă prin detașarea bulelor mici de pe suprafața sa în celulă și încorporarea vacuolelor din interiorul celulei în membrană. Astfel, există un flux constant de elemente membranare în celulă: din membrana plasmatică în citoplasmă (endocitoza)și fluxul structurilor membranare de la citoplasmă la suprafața celulei (exocitoză).În turnover-ul membranei, rolul principal este jucat de sistemul de vacuole membranare ale complexului Golgi.
4. Funcțiile membranei plasmatice. Mecanisme de transport al substantelor prin plasmalema. Funcția de receptor a plasmalemei
Membrana plasmatică îndeplinește o serie de funcții importante:
1) Barieră.Funcția de barieră a membranei plasmatice este de alimitarea difuziei libere a substantelor de la celula la celula, prevenindscurgerea rotativă a conținutului celulelor solubile în apă. Dar de atuncicelula ta trebuie să primească nutrienții necesari, tuîmparte produsele finale ale metabolismului, reglează intracelularDacă concentrația de ioni este mare, atunci s-au format mecanisme speciale pentru transferul de substanțe prin membrana celulară.
2) Transport.Funcția de transport include asigurând intrarea și ieșirea diferitelor substanțe în și din celulă. O proprietate importantă a membranei este permeabilitate selectivă, sau semi-permeabilitate. Trece cu ușurință apa și soluțiile de apăgaze și respinge moleculele polare precum glucoza sau aminoacizi.
Există mai multe mecanisme pentru transportul substanțelor prin membrană:
transport pasiv;
transport activ;
transport în ambalaj cu membrană.
Transport pasiv. Difuzie -aceasta este mișcarea particulelor din mediu, care duce la transferul de energiesubstanțe dintr-o zonă în care concentrația sa este mare până la o zonă cu concentrație scăzutăție. În timpul transportului prin difuzie, membrana funcționează ca o barieră osmotică. Viteza de difuzie depinde de mărimemolecule și solubilitatea relativă a acestora în grăsimi. Cu cât mai puține orimăsurile de molecule și cu cât sunt mai solubile în grăsimi (lipofile), cu atât se vor deplasa mai repede prin stratul dublu lipidic.Difuzia poate fi neutru(transferul neîncărcatmolecule) şi ușoară(cu ajutorul proteinelor specialepurtători). Rata difuziei facilitate este mai mare decât cea a difuziei neutre.Penetrare maximaApa are capacitatea de acum moleculele sale sunt mici și neîncărcate. Difuzia apei prin celulemembrana se numeste osmo somnSe presupune că în celulemembrana pentru penetrareapa si unii ioni suntexistă „pori” speciali. Numărul loreste mic, iar diametrul esteaproximativ 0,3-0,8 nm. Se difuzează cel mai repede prin membrană bine, ușor solubil în lipide dublu strat al unei molecule, cum ar fi O și molecule polare neîncărcateleșii cu diametru mic (SO, mo chevina).
Transferul de molecule polare (cuzaharuri, aminoacizi), în specialfabricat folosind transport special cu membranăproteinele se numesc difuzie facilitată. Se găsesc astfel de proteinegăsit în toate tipurile de membrane biologice și fiecare specific Această proteină este concepută pentru a transporta molecule dintr-o anumită clasă sa. Proteinele de transport sunt transmembranare; lanțul lor polipeptidic traversează de mai multe ori stratul lipidic, formându-se Are treceri prin intermediul. Aceasta asigură transferul de specificsubstanțe prin membrană fără contact direct cu aceasta.Există două clase principale de proteine de transport: proteine- transportatori (transportatori) Și formarea canalelor proteine (albecanale ki). Proteinele purtătoare transportă molecule prin membrană, mai întâi schimbându-le configurația. Proteinele care formează canale formează membrane umplute porii de apă. Când porii sunt deschiși, molecule de substanțe specifice(de obicei, ioni anorganici de mărime și sarcină adecvate) trec prin ele. Dacă molecula substanței transportate nu are nicio sarcină, atunci direcția de transport este determinată de gradientul de concentrație. Dacă molecula este încărcată, atunci transportul ei, pe lângă gradient, depinde centralizare, sarcina electrică a membranei (membranăpotenţial). Partea interioară a plasmalemei este de obicei încărcată din negativ în raport cu exteriorul. Potențialul de membrană facilitează pătrunderea ionilor încărcați pozitiv în celulă și împiedică trecerea ionilor încărcați negativ.
Transport activ. Transportul activ este mișcarea substanțelor împotriva unui gradient electrochimic. Este întotdeauna realizată de proteinele transhamali şi strâns înrudiţi zan cu sursa de energiegii. În transferul de proteine sunt parcele obligatoriu de transportsubstanta titrata. Cu cât mai multe astfel de studii tkov contactează chestiacu atât rata este mai marecresterea transportului. Transferul selectiv al unei substanțe se numește uniport. Se efectuează transferul mai multor substanțe Kotran sisteme sportive. Dacă transferul merge într-o singură direcție -Acest simport, dacă în sens opus - antiport. Asa de,de exemplu, glucoza este transferată din fluidul extracelular în celulă uniport. Transferul de glucoză și Na 4 din cavitatea intestinală sautubii renali, respectiv, în celulele intestinale sau în sânge se efectuează simportal, iar transferul de C1~ și HCO este antiportor. Se presupune că în timpul transferului apar modificări conformaționale reversibile. modificări ale transportorului, care permite mișcarea substanțelor conectate la acesta.
Un exemplu de proteină purtătoare utilizată pentru transportsubstanţe, energia eliberată în timpul hidrolizei ATP esteN / A + -K + pompă, găsite în membrana plasmatică a tuturor celulelor. Na+-K pompa functioneaza pe principiul antiport, pompare vaya Na „din celulă și K t în celulă împotriva lor electrochimică gradiente. Gradient Na+ creeaza presiune osmotica, mentine volumul celular si asigura transportul zaharurilor si aminoacizilornoacizi Functionarea acestei pompe consuma o treime din toata energia necesara functionarii celulelor.La studierea mecanismului de acţiune Na+ - K+ pompa a fost instalataSe arată că este o enzimă ATPază și o proteină transmembranară. proteine integrale. In prezenta Na+ și ATP sub influența ATP-Fosfatul terminal este separat de ATP și adăugat la restulacid aspartic pe o moleculă de ATPază. Molecula de ATPaza fosforilează, își schimbă configurația și Na+ este eliminat din celule. În urma retragerii N / A K" este întotdeauna transportat din celulă în celulă. Pentru a face acest lucru, fosfatul atașat anterior este scindat din ATPază în prezența K. Enzima este defosforilată, își restabilește configurația și K 1 este "pompat" în celulă.
ATPaza este formată din două subunități, mari și mici.Subunitatea mare este formată din mii de reziduuri de aminoacizi,traversând stratul dublu de mai multe ori. Are un catalitic activitate și poate fi fosforilat și defosforizat reversibila fi realizat. Subunitate mare pe partea citoplasmaticănu are zone pentru legare Na+ și ATP, iar în exterior -situsuri de legare pentru K+ și ouabaină. Subunitatea mică esteglicoproteina și funcția acesteia nu este încă cunoscută.
Na+-K pompa are efect electrogen. El scoate treiion încărcat pozitiv Na f din cușcă și aduce douăion K Ca urmare, un curent trece prin membrană, formând un electrodpotențial ric cu o valoare negativă în interiorul celulei în raport cu suprafața ei exterioară. Na"-K+ pompa reglează volumul celular, controlează concentrația de substanțeîn interiorul celulei, menține presiunea osmotică, participă la crearea potențialului de membrană.
Transport in ambalaj membrana. Transferul de macromolecule (proteine, acizi nucleici) prin membranălot, polizaharide, lipoproteine) și alte particule sunt realizate prin formarea și fuziunea secvențială a înconjurat.vezicule legate de membrană (vezicule). Procesul de transport vezicularse desfășoară în două etape. La inceputmembrana veziculelor si plasmalemalipiți împreună și apoi îmbinați.Pentru ca etapa 2 să aibă loc este necesarMi-aș dori să fiți molecule de apăsunt aglomerate prin interacțiuni duble straturi lipidice, care se apropie la o distanță de 1-5 nm. Contează Xia că acest proces este activatspecial proteine de fuziune(Ei izolat până acum doar de viruși). Transportul vezicular arecaracteristică importantă - macromolecule absorbite sau secretate,situat în bule, de obicei nuse amestecă cu alți macromolculi sau organite ale celulei. Pu cosurile se pot îmbina cu specificul membrane chimice, care asigurăfacilitează schimbul de macromolecule întreîntre spaţiul extracelular şicontinutul celulei. De asemeneamacromoleculele sunt transferate dintr-un compartiment celular în altul.
Transportul macromoleculelor și particulelor în celulă se numește endo citoza.În acest caz, substanțele transportate sunt învelitede membrana plasmatică se formează o veziculă (vacuola), carecare se deplasează în interiorul celulei. În funcție de dimensiunea imaginiiformând vezicule, există două tipuri de endocitoză - pinocitoza si fagocitoza.
Pinocitozaasigură absorbția lichidului și dizolvatsubstanțe sub formă de bule mici ( d =150 nm). fagocitoza -aceasta este absorbția particulelor mari, a microorganismelorapel sau fragmente de organite, celule. În acest caz se formeazăexistă vezicule mari, fagozomi sau vacuole ( d -250 nm sau mai mult). U funcția fagocitară a protozoarelor – formă de nutriție. La mamifere, funcția fagocitară este îndeplinită de macrofage șirofils, care protejează organismul de infecții prin absorbția microbilor invadatori. Macrofagele sunt, de asemenea, implicate în reciclarecelule vechi sau deteriorate și resturile acestora (în organismmacrofagele umane consumă zilnic peste 100 de erite vechirocite). Fagocitoza începe numai atunci când particulele înghițitese leagă de suprafața fagocitelor și activează specializatny celule receptore. Legarea particulelor de substanțe specificereceptorii membranari determină formarea pseudopodiilor, careEle învăluie particula și, fuzionandu-se la margini, formează o bulă -fagozom.Are loc formarea unui fagozom și fagocitoza în sinese mișcă numai dacă, în timpul procesului de învelire, particulacontact constant cu receptorii plasmalemei, ca și cum ar „stagna” fulger fulgerător”.
O parte semnificativă a materialului absorbit de celulă prin intermediul endocitoză, își încheie călătoria în lizozomi. Particule mari incluseașteaptă cu nerăbdare fagozomi, care apoi fuzionează cu lizozomii şi formează fagolizozomi. Lichid și macromolecule absorbite în timpulpinocitoza, sunt inițial transferate la endozomi, care suntse fuzionează cu lizozomii formând endolizozomi. sunt prezent diverse enzime hidrolitice prezente rapid în lizozomidistruge macromoleculele. Produse de hidroliză (aminoaciziloturi, zaharuri, nucleotide) sunt transportate de la lizozomi la citosol, unde sunt folosite de celulă. Majoritatea componentelor membranei veziculele endocitare din fagozomi și endozomi revin prin exocitoză la membrana plasmatică și sunt redistribuite acolosunt lizate. Principala semnificație biologică a endocitozei este este posibil să se obţină blocuri de construcţie datorită intracelulare digestia macromoleculelor din lizozomi.
Absorbția substanțelor în celulele eucariote începe înzonele specializate ale membranei plasmatice, așa-numitelenoi suntem X gropi mărginite.În micrografiile electronicegropile arată ca invaginări ale membranei plasmatice, citoplasmeia cărui latură mată este acoperită cu un strat fibros. Strat caar mărgini cu micile gropi ale pieței Malemele. Gropile ocupă aproximativ 2% vol.suprafața membranei celularenoi eucariotele. Într-un minut gropile cresc, sapă din ce în ce mai adânc Xia, sunt atrași în celulă și apoi, se îngustează la bază, se desprinde,formând bule mărginite.S-a stabilit ca din piatamembrana mat fibroblastTovarăș în decurs de un minutse toarnă aproximativ un sfertmembrane sub formă de PU mărginite Zyrkov. Bulele dispar repede granița lor și dobândește o calecapacitatea de a fuziona cu lizozomul.
Endocitoza poate fi nespecific(constitutiv)Și specific(receptor).La endocitoză nespecifică celula preia şiabsoarbe substanțe complet străine de el, de exemplu, particule de funingine,coloranți. În primul rând, particulele sunt depuse pe glicocalix. plasmaleme. Sunt deosebit de bine depuse (adsorbite) de grupuri de proteine încărcate pozitiv, deoarece glicocalixul poartă sarcina negativa. Apoi morfologia celulei se schimbămembranelor. Se poate scufunda fie, formând invaginări(invaginări) sau, dimpotrivă, să formeze excrescențe, care par să se plieze, separând volumele mici mediu lichid. Formarea invaginațiilor este mai tipică pentru celulele epiteliale intestinale, amibe și excrescențe - pentru fagocite și fibroblaste. Aceste procese pot fi blocate cu inhibitorirespiraţie. Veziculele rezultate sunt endozomi primari și se pot scurge schimbând între ele, crescând în dimensiune. Mai târziu se vor conecta interacționează cu lizozomii, transformându-se într-un endolizozom - digestiv vacuola noua. Intensitatea pinocitozei nespecifice în fază lichidă până ladestul de inalt. Macrofagele formează până la 125, iar celulele epiteliale sunt subțiriintestinele până la o mie de pinos pe minut. Abundența pinozomilor duce la faptul că plasmalema este cheltuită rapid pentru formarea multorde vacuole mici. Restaurarea membranei este destul de rapidătro în timpul reciclării în timpul exocitozei datorită întoarcerii vacuole şi integrarea lor în plasmalemă. Macrofagele au toată plasmăMembrana chimică se înlocuiește în 30 de minute, iar în fibroblaste în 2 ore.
Mod mai eficient de absorbție din lichidul extracelularmacromoleculele specifice osoase este specific ro docitoză(mediat de receptor). În același timp, macromoleculese leagă de receptorii complementari de la suprafațăcelulele se acumulează în groapa mărginită, iar apoi, formând un endozom, sunt scufundate în citosol. Endocitoza receptorului asigură acumularea de macromolecule specifice la receptorul său.Molecule care se leagă la suprafața plasmalemei cu receptoritorus sunt numite liganzi. Folosind receptorul endocitoza în multe celule animale are loc absorbțiacolesterol din extracelular mediu inconjurator.
Membrana plasmatică participă la eliminarea substanțelor din celulă (exocitoză). În acest caz, vacuolele se apropie de plasmalemă. În punctele de contact, membrana plasmatică și membrana vacuolei se îmbină și conținutul vacuolei intră în mediul înconjurător.La unele protozoare, locurile de pe membrana celulară pentru exocitoză sunt predeterminate. Deci, în membrana plasmatică Unii ciliati ciliati au anumite zone cu aranjarea corecta a globulelor mari de proteine integrale. Umucochisturile și tricochisturile ciliatelor sunt complet gata pentru secreție; în partea superioară a plasmalemei există o margine de globule integrale.proteine. Aceste zone ale membranei mucochiste și tricochisturi sunt adiacenteaderă la suprafața celulei.La neutrofile se observă un fel de exocitoză. Suntpoate, în anumite condiții, să fie eliberat în mediufă-ți lizozomii. În unele cazuri, se formează mici excrescențe ale plasmalemei care conțin lizozomi, care apoi se desprind și se mută în mediu. În alte cazuri, se observă invaginarea plasmalemei adânc în celulă și captarea acesteia de lizozomi, localizată situat departe de suprafața celulei.
Procesele de endocitoză și exocitoză sunt efectuate cu participarea unui sistem de componente fibrilare ale citoplasmei asociate cu plasmalema.
Funcția de receptor a plasmalemei. Acesta este unul una dintre cele principale, universală pentru toate celulele, este refuncția de receptor a plasmalemei. Ea definește interacțiuneacelule între ele și cu mediul extern.
Întreaga varietate de interacțiuni intercelulare informaționale poate fi reprezentată schematic ca un lanț de secvențialereacții semnal-receptor-mesager secund-răspuns (concept semnal-răspuns).Semnalele transmit informații de la celulă la celulămolecule care sunt produse în anumite celule și specialeinfluențează fizic alte celule sensibile la semnal (celule) sheni). Molecula semnal - intermediar primar legând interacționează cu receptorii localizați pe celulele țintă, reacționează transmiterea numai la anumite semnale. Molecule semnal - liganzi- se potrivește receptorului său ca pe o cheie a unui încuietor. Ligand-pentru receptorii membranari (receptorii plasmalema) suntmolecule hidrofile, hormoni peptidici, neuromedia- tori, citokine, anticorpi, iar pentru receptorii nucleari - grăsime Molecule romane, hormoni steroizi și tiroidieni, vitamina DCa receptori deasupraproteina poate acționa ca o celulămembrane sau elemente de glicocalixca - polizaharide și glicoproteine.Se crede că sunt sensibili lazone, împrăștiatesan pe suprafata celulei sau cubranele în zone mici. Da, pesuprafața celulelor procarioteiar celulele animale există limitenumăr limitat de locuri cu care potleagă particulele virale. Eu eujură proteine (transportatori și canaly) recunoaște, interacționează și transferătransportă doar anumite substanțe.Receptorii celulari sunt implicați întransmiterea semnalelor de la suprafața celulei în ea.Diversitate și specificitateșanț de receptori de pe suprafața celuleiconduce la crearea unui sistem foarte complexavem markeri care ne permit să distingemcelulele tale de la alții. Celule similareinteracționează între ele, suprafețele lor se pot lipi între ele (conjugareprotozoare, formarea țesuturilor în organisme pluricelulare). Nu percep celulelemarkeri comuni, precum și cei care diferă înborul markerilor determinanțiagățați sau respingeți.La formarea complexului receptor-ligand, acestea sunt activateproteine transmembranare: proteină traductoare, proteină amplificatoare.Ca urmare, receptorul își schimbă conformația și interacțiuneaexistă cu precursorul celui de-al doilea mesager situat în celulă ka - mesager.Mesagerii pot fi calciu ionizat, fosfolipidepentru C, adenilat ciclaza, guanilat ciclaza. Sub influența mesageruluienzimele implicate în sinteza sunt activate monofosfați ciclici - AMP sau GMF. Acestea din urmă schimbă activulprezența a două tipuri de enzime protein kinaze în citoplasma celulară, ducând la fosforilarea a numeroase proteine intracelulare.
Cea mai frecventă este formarea cAMP, sub influența cocare crește secreția unui număr de hormoni - tiroxină, cortizon, progesteron, crește descompunerea glicogenului în ficat și mușchi,ritmul și forța cardiacă, osteodistrucție, invers absorbția apei în tubii nefronici.
Activitatea sistemului de adenil-ciclază este foarte mare - sinteza cAMP duce la o creștere a zece miimi a semnalului.
Sub influența cGMP, crește secreția de insulină de către pancreas, histamina de către mastocitele și serotonina de către trom.bocite, țesutul muscular neted se contractă.
În multe cazuri, când se formează un complex receptor-ligandexistă o modificare a potențialului membranei, care, la rândul său, duce la o modificare a permeabilității plasmalemei și a metabolismului.unele procese din celulă.
Receptorii specifici sunt localizați pe membrana plasmatică tori care răspund la factori fizici. Astfel, în bacteriile fotosintetice, clorofilele sunt localizate pe suprafața celulei,receptiv la lumină. La animalele fotosensibile din plasmăMembrana cerebrală conține un întreg sistem de proteine fogoreceptoare -rodopsine, cu ajutorul cărora stimulul luminos se transformă transformat într-un semnal chimic și apoi într-un impuls electric.
3. Contacte intercelulare
În organismele animale multicelulare, plasmalema participă la formare conexiuni intercelulare, oferind interacțiuni intercelulare. Există mai multe tipuri de astfel de structuri.
§ Contact simplu.Contactul simplu are loc între majoritatea celulelor adiacente de diferite origini. Reprezintă convergența membranelor plasmatice ale celulelor învecinate la o distanță de 15-20 nm. În acest caz, are loc interacțiunea straturilor de glicocalix ale celulelor învecinate.
§ Contact strâns (închis). Cu această legătură, straturile exterioare ale celor două membrane plasmatice sunt cât mai apropiate. Apropierea este atât de apropiată încât este ca și cum secțiunile plasmalemei a două celule învecinate se unesc. Fuziunea membranei nu are loc pe întreaga zonă de contact strâns, ci reprezintă o serie de abordări punctiforme ale membranelor. Rolul joncțiunii strânse este de a conecta mecanic celulele între ele. Această zonă este impermeabilă la macromolecule și ioni și, prin urmare, închide și delimitează golurile intercelulare (și odată cu acestea mediul intern al organismului) de mediul extern.
§ Punct de coeziune sau desmozom. Desmozomul este o zonă mică cu un diametru de până la 0,5 microni. În zona desmozomului de pe partea citoplasmatică există o zonă de fibrile subțiri. Rolul funcțional al desmozomilor este în principal comunicarea mecanică între celule.
§ Intersecție sau nexus. Cu acest tip de contact, membranele plasmatice ale celulelor vecine sunt separate printr-un interval de 2-3 nm pe o distanță de 0,5-3 µm. Structura membranelor plasmatice conține complexe proteice speciale (conexoni). Un conexon de pe membrana plasmatică a unei celule este exact opus unui conexon de pe membrana plasmatică a unei celule adiacente. Ca rezultat, se formează un canal de la o celulă la alta. Conexonii se pot contracta, modificând diametrul canalului intern și, prin urmare, participă la reglarea transportului de molecule între celule. Acest tip de conexiune se găsește în toate grupele de țesuturi. Rolul funcțional al joncțiunii gap este de a transporta ionii și moleculele mici de la celulă la celulă. Astfel, în mușchiul cardiac, excitația, care se bazează pe procesul de modificare a permeabilității ionice, este transmisă de la celulă la celulă prin nexus.
§ Contact sinaptic sau sinapsa. Sinapsele sunt zone de contact între două celule specializate pentru transmiterea unilaterală a excitației sau inhibiției de la un element la altul. Acest tip de conexiune este caracteristic țesutului nervos și are loc atât între doi neuroni, cât și între un neuron și un alt element. Membranele acestor celule sunt separate printr-un spațiu intercelular - o despicatură sinaptică de aproximativ 20-30 nm lățime. Membrana din zona de contact sinaptic a unei celule se numește presinaptic, cealaltă - postsinaptic. În apropierea membranei presinaptice, sunt detectate un număr mare de vacuole mici (vezicule sinaptice) care conțin emițătorul. În momentul trecerii unui impuls nervos, veziculele sinaptice eliberează transmițătorul în fanta sinaptică. Mediatorul interacționează cu situsurile receptorilor membranei postsinaptice, ceea ce duce în cele din urmă la transmiterea unui impuls nervos. Pe lângă transmiterea impulsurilor nervoase, sinapsele asigură o legătură rigidă între suprafețele a două celule care interacționează.
§ Plasmodesmate.Acest tip de comunicare intercelulară se găsește la plante. Plasmodesmatele sunt canale tubulare subțiri care conectează două celule adiacente. Diametrul acestor canale este de obicei de 40-50 nm. Plasmodesmele trec prin peretele celular care separă celulele. În celulele tinere, numărul de plasmodesmate poate fi foarte mare (până la 1000 per celulă). Pe măsură ce celulele îmbătrânesc, numărul lor scade din cauza rupturilor pe măsură ce grosimea peretelui celular crește. Rolul funcțional al plasmodesmelor este de a asigura circulația intercelulară a soluțiilor care conțin nutrienți, ioni și alți compuși. Prin plasmodesmate, celulele sunt infectate cu virusuri vegetale.
Structuri specializate ale membranei plasmatice
Plasmalema multor celule animale formează excrescențe ale diferitelor structuri (microvili, cili, flageli). Cel mai adesea se găsește pe suprafața multor celule animale microvilozități. Aceste excrescențe ale citoplasmei, limitate de plasmalemă, au forma unui cilindru cu vârful rotunjit. Microvilozitățile sunt caracteristice celulelor epiteliale, dar se găsesc și în celulele altor țesuturi. Diametrul microvilozităților este de aproximativ 100 nm. Numărul și lungimea lor variază între diferitele tipuri de celule. Semnificația microvilozităților este creșterea semnificativă a suprafeței celulare. Acest lucru este deosebit de important pentru celulele implicate în absorbție. Astfel, în epiteliul intestinal există până la 2x10 8 microvilli la 1 mm 2 de suprafaţă.
Membrana plasmatică ocupă o poziție specială, deoarece limitează celula din exterior și este direct conectată la mediul extracelular. Are o grosime de aproximativ 10 nm și este cea mai groasă dintre membranele celulare. Componentele principale sunt proteinele (mai mult de 60%), lipidele (aproximativ 40%) și carbohidrații (aproximativ 1%). Ca toate celelalte membrane celulare, este sintetizat în canalele ER.
Funcțiile plasmalemei.
Transport.
Membrana plasmatică este semipermeabilă, adică. prin ea trec selectiv diferite molecule cu viteze diferite. Există două moduri de a transporta substanțe prin membrană: transport pasiv și activ.
Transport pasiv. Transportul pasiv sau difuzia nu necesită energie. Moleculele neîncărcate difuzează de-a lungul unui gradient de concentrație; transportul moleculelor încărcate depinde de gradientul de concentrație al protonilor de hidrogen și de diferența de potențial transmembranar, care se combină pentru a forma un gradient de protoni electrochimic. De regulă, suprafața citoplasmatică interioară a membranei poartă o sarcină negativă, care facilitează pătrunderea ionilor încărcați pozitiv în celulă. Există două tipuri de difuzie: simplă și facilitată.
Difuzia simplă este tipică pentru molecule neutre mici (H 2 O, CO 2, O 2), precum și pentru substanțele organice hidrofobe cu greutate moleculară mică. Aceste molecule pot trece fără nicio interacțiune cu proteinele membranei prin porii sau canalele membranei atâta timp cât se menține gradientul de concentrație.
Difuzia facilitată este caracteristică moleculelor hidrofile care sunt transportate prin membrană și de-a lungul unui gradient de concentrație, dar cu ajutorul proteinelor speciale de transport membranare conform principiului uniport.
Difuzia facilitată este foarte selectivă, deoarece proteina purtătoare are un centru de legare complementar substanței transportate, iar transferul este însoțit de modificări conformaționale ale proteinei. Unul dintre mecanismele posibile de difuzie facilitată este următorul: o proteină de transport (translocaza) leagă o substanță, apoi se apropie de partea opusă a membranei, eliberează această substanță, își ia conformația inițială și este din nou gata să îndeplinească funcția de transport. Se știe puțin despre cum se mișcă proteina în sine. Un alt mecanism posibil de transport implică participarea mai multor proteine transportoare. În acest caz, compusul legat inițial în sine se deplasează de la o proteină la alta, legându-se secvenţial cu una sau cealaltă proteină până când ajunge pe partea opusă a membranei.
Transport activ. Un astfel de transport are loc atunci când transportul are loc în raport cu un gradient de concentrație. Necesită consum de energie de către celulă. Transportul activ servește la acumularea de substanțe în interiorul celulei. Sursa de energie este adesea ATP. Pentru transportul activ, pe lângă sursa de energie, este necesară participarea proteinelor membranare. Unul dintre sistemele de transport active din celulele animale este responsabil pentru transportul ionilor de Na și K + prin membrana celulară. Acest sistem este numit N / A + - Pompa K*. Este responsabil pentru menținerea compoziției mediului intracelular, în care concentrația de ioni K + este mai mare decât ionii Na *.
Gradientul de concentrație al ambilor ioni este menținut prin transferul de K + în celulă și Na + afară. Ambele transporturi au loc împotriva gradientului de concentrație. Această distribuție a ionilor determină conținutul de apă din celule, excitabilitatea celulelor nervoase și a celulelor musculare și alte proprietăți ale celulelor normale. Pompa Na + -K + - este o proteină - transport ATPaza. Molecula acestei enzime este un oligomer și pătrunde în membrană. În timpul unui ciclu complet de funcționare a pompei, 3 ioni Na + sunt transferați de la celulă la substanța intercelulară, iar 2 ioni K + sunt transferați în direcția opusă, folosind energia moleculei ATP. Există sisteme de transport pentru transferul ionilor de calciu (Ca 2+ -ATPaze), pompe de protoni (H + -ATPaze), etc.
Se numește transferul activ al unei substanțe printr-o membrană, realizat datorită energiei gradientului de concentrație al unei alte substanțe simport. ATPaza de transport în acest caz are centre de legare pentru ambele substanțe. Antiport este mișcarea unei substanțe față de gradientul său de concentrație. În acest caz, o altă substanță se mișcă în direcția opusă de-a lungul gradientului său de concentrație. Simportul și antiportul (cotransportul) pot apărea în timpul absorbției aminoacizilor din intestin și reabsorbției glucozei din urina primară, folosind energia gradientului de concentrație a ionilor Na + creat de Na + , K + -ATPaza.
Încă două tipuri de transport sunt endocitoza și exocitoza.
Endocitoza- captarea particulelor mari de către celulă. Există mai multe moduri de endocitoză: pinocitoză și fagocitoză. De obicei sub pinocitozaînțelegeți absorbția particulelor coloidale lichide de către celulă, sub fagocitoză- captarea corpusculilor (particule mai dense si mai mari pana la alte celule). Mecanismul de pino- și fagocitoză este diferit.
În general, intrarea particulelor solide sau a picăturilor de lichid într-o celulă din exterior se numește heterofagie. Acest proces este cel mai răspândit la protozoare, dar este foarte important și la om (precum și la alte mamifere). Heterofagia joacă un rol semnificativ în apărarea organismului (neutrofile segmentate - granulocite; macrofagocite), restructurarea țesutului osos (osteoclaste), formarea tiroxinei de către foliculii tiroidieni, reabsorbția proteinelor și a altor macromolecule în nefronul proximal și alte proceselor.
Pinocitoza.
Pentru ca moleculele externe să pătrundă în celulă, acestea trebuie mai întâi legate de receptorii glicocalix (un set de molecule asociate cu proteinele membranei de suprafață) (Fig.).
La locul unei astfel de legături, moleculele de proteină clatrină se găsesc sub plasmalemă. Membrana plasmatică, împreună cu moleculele atașate din exterior și acoperite de clatrină pe partea citoplasmei, începe să se invagineze. Invaginația devine mai profundă, marginile ei se apropie și apoi se închid. Ca rezultat, o bulă care conține molecule prinse se desprinde de plasmalemă. Clathrin pe suprafața sa arată ca o margine neuniformă pe micrografiile electronice, motiv pentru care astfel de bule sunt numite marginate.
Clathrin previne atașarea veziculelor de membranele intracelulare. Prin urmare, veziculele mărginite pot fi transportate liber în celulă tocmai în acele zone ale citoplasmei în care conținutul lor ar trebui utilizat. Acesta este modul în care hormonii steroizi, în special, sunt eliberați în nucleu. Cu toate acestea, de obicei veziculele mărginite își dispar granița la scurt timp după ce s-au desprins de plasmalemă. Clatrina este transportată la plasmalemă și poate participa din nou la reacțiile de endocitoză.
În apropierea suprafeței celulare în citoplasmă există mai multe vezicule permanente - endozomi. Veziculele mărginite elimină clatrina și fuzionează cu endozomii, crescând astfel volumul și suprafața endozomilor. Apoi, partea în exces a endozomilor este divizată sub forma unei noi vezicule, în care nu există substanțe care pătrund în celulă; acestea rămân în endozom. Noua veziculă călătorește la suprafața celulei și fuzionează cu membrana. Ca urmare, pierderea plasmalemei, care a avut loc atunci când vezicula mărginită a fost detașată, este restabilită, iar receptorii săi revin și ei la plasmalemă.
Endozomii sunt scufundați în citoplasmă și se îmbină cu membranele lizozomului. Substanțele care intră în interiorul unui astfel de lizozom secundar suferă diferite transformări biochimice. La finalizarea procesului, membrana lizozomului se poate dezintegra în fragmente, iar produsele de descompunere și conținutul lizozomului devin disponibile pentru reacții metabolice intracelulare. De exemplu, aminoacizii sunt legați de ARNt și eliberați la ribozomi, iar glucoza poate intra în complexul Golgi sau în tubii ER agranulare.
Deși endozomii nu au o margine de clatrină, nu toți fuzionează cu lizozomi. Unele dintre ele sunt direcționate de la o suprafață celulară la alta (dacă celulele formează un strat epitelial). Acolo, membrana endozomală se contopește cu plasmalema și conținutul este îndepărtat în exterior. Ca urmare, substanțele sunt transferate prin celulă dintr-un mediu în altul fără modificări. Acest proces se numește transcitoza. Moleculele de proteine, în special imunoglobulinele, pot fi de asemenea transferate prin transcitoză.
Fagocitoză.
Dacă o particulă mare are pe suprafața sa grupe moleculare care pot fi recunoscute de receptorii celulari, se leagă. Nu este întotdeauna cazul ca particulele străine în sine să posede astfel de grupuri. Cu toate acestea, atunci când intră în organism, sunt înconjurate de molecule de imunoglobuline (opsonine), care se găsesc întotdeauna atât în sânge, cât și în mediul intercelular. Imunoglobulinele sunt întotdeauna recunoscute de celulele fagocitare.
După ce opsoninele care acoperă particulele străine s-au legat de receptorii fagocitari, complexul său de suprafață este activat. Microfilamentele de actină încep să interacționeze cu miozina, iar configurația suprafeței celulare se modifică. Excrescențe ale citoplasmei fagocitelor se extind în jurul particulei. Acopera suprafața particulei și se combină deasupra acesteia. Straturile exterioare ale excrescentelor se îmbină, închizând suprafața celulei.
Plăcile adânci de excrescențe formează o membrană în jurul particulei absorbite - a fagozom. Fagozomul fuzionează cu lizozomii, rezultând complexul lor - heterolizozom (heterozom, sau fagolizozom).În ea, are loc liza componentelor capturate ale particulei. Unii dintre produșii de liză sunt îndepărtați din heterozom și utilizați de celulă, în timp ce unii nu pot fi susceptibili de acțiunea enzimelor lizozomale. Aceste reziduuri formează corpuri reziduale.
Potenţial toate celulele au capacitatea de a fagocita, dar în organism doar câteva sunt specializate în această direcţie. Acestea sunt leucocite neutrofile și macrofage.
exocitoză.
Aceasta este eliminarea substanțelor din celulă. În primul rând, compușii moleculari mari sunt segregați în complexul Golgi sub formă de vezicule de transport. Acestea din urmă, cu participarea microtubulilor, sunt direcționate către suprafața celulei. Membrana veziculei este înglobată în plasmalemă, iar conținutul veziculei apare în afara celulei (Fig.) Vezicula poate fuziona cu plasmalema fără semnale suplimentare. Această exocitoză se numește constitutiv. Acesta este modul în care majoritatea produselor propriului metabolism sunt îndepărtate din celulă. O serie de celule, cu toate acestea, sunt concepute pentru a sintetiza compuși speciali - secreții care sunt utilizate în alte părți ale corpului. Pentru ca vezicula de transport cu secreția să se îmbine cu plasmalema, sunt necesare semnale din exterior. Abia atunci va avea loc fuziunea și secretul va fi dezvăluit. Această exocitoză se numește reglabil. Se numesc molecule semnal care promovează eliminarea secrețiilor liberine (factori de eliberare),și cei care împiedică excreția - statine.
Funcțiile receptorului.
Ele sunt furnizate în principal de glicoproteinele situate pe suprafața plasmalemei și capabile să se lege de liganzii lor. Ligandul corespunde receptorului său ca o cheie a unui lacăt. Legarea unui ligand la un receptor determină o modificare a conformației polipeptidei. Odată cu această modificare a proteinei transmembranare, se stabilește comunicarea între mediul extracelular și cel intracelular.
Tipuri de receptori.
Receptori cuplati la canalele ionice proteice. Ele interacționează cu o moleculă de semnalizare, care deschide sau închide temporar canalul pentru trecerea ionilor. (De exemplu, receptorul pentru neurotransmițătorul acetilcolină este o proteină formată din 5 subunități care formează un canal ionic. În absența acetilcolinei, canalul este închis, iar după atașare se deschide și permite trecerea ionilor de sodiu).
Receptorii catalitici. Ele constau dintr-o parte extracelulară (receptorul însuși) și o parte citoplasmatică intracelulară, care funcționează ca o enzimă prolinkază (de exemplu, receptorii hormonului de creștere).
Receptorii cuplați cu proteina G. Acestea sunt proteine transmembranare formate dintr-un receptor care interacționează cu ligand și o proteină G (proteină reglatoare legată de guanozin trifosfat) care transmite un semnal către o enzimă legată de membrană (adenilat ciclază) sau un canal ionic. Ca rezultat, AMP ciclic sau ionii de calciu sunt activați. (Așa funcționează sistemul de adenil ciclază. De exemplu, în celulele hepatice există un receptor pentru hormonul insulină. Partea supracelulară a receptorului se leagă de insulină. Acest lucru determină activarea părții intracelulare - enzima adenilat ciclază. Se sintetizează AMP ciclic din ATP, care reglează viteza diferitelor procese intracelulare, determinând activarea sau inhibarea acelor sau a altor enzime metabolice).
Receptorii care percep factorii fizici. De exemplu, proteina fotoreceptor rodopsina. Când lumina este absorbită, aceasta își schimbă conformația și excită un impuls nervos.
Celulă– o unitate elementară a unui sistem viu. Diverse structuri ale unei celule vii care sunt responsabile pentru îndeplinirea unei anumite funcții sunt numite organele, ca și organele unui întreg organism. Funcțiile specifice în celulă sunt distribuite între organele, structuri intracelulare care au o anumită formă, precum nucleul celular, mitocondriile etc.
Structuri celulare:
Citoplasma. O parte esențială a celulei, închisă între membrana plasmatică și nucleu. Citosol este o soluție apoasă vâscoasă de diferite săruri și substanțe organice, impregnată cu un sistem de fire proteice - citoschelete. Cele mai multe procese chimice și fiziologice ale celulei au loc în citoplasmă. Structura: Citosol, citoschelet. Funcții: include diverse organite, mediul celular intern
Membrană plasmatică. Fiecare celulă de animale, plante, este limitată de mediu sau de alte celule de o membrană plasmatică. Grosimea acestei membrane este atât de mică (aproximativ 10 nm) încât poate fi văzută doar cu un microscop electronic.

Lipidele formează un strat dublu în membrană, iar proteinele pătrund în toată grosimea acesteia, sunt scufundate la diferite adâncimi în stratul lipidic sau sunt situate pe suprafețele exterioare și interioare ale membranei. Structura membranelor tuturor celorlalte organite este similară cu membrana plasmatică. Structura: strat dublu de lipide, proteine, carbohidrati. Funcții: restrângerea, păstrarea formei celulelor, protecția împotriva deteriorării, reglator al aportului și eliminarea substanțelor.
Lizozomi. Lizozomii sunt organite legate de membrană. Au o formă ovală și un diametru de 0,5 microni. Acestea conțin un set de enzime care distrug substanțele organice. Membrana lizozomilor este foarte puternică și împiedică pătrunderea propriilor enzime în citoplasma celulei, dar dacă lizozomul este deteriorat de orice influențe externe, atunci întreaga celulă sau o parte a acesteia este distrusă.
Lizozomii se găsesc în toate celulele plantelor, animalelor și ciupercilor.
Prin digerarea diferitelor particule organice, lizozomii furnizează „materii prime” suplimentare pentru procesele chimice și energetice din celulă. Când celulele sunt înfometate, lizozomii digeră unele organele fără a ucide celula. Această digestie parțială oferă celulei minimul necesar de nutrienți pentru o perioadă de timp. Uneori, lizozomii digeră celule întregi și grupuri de celule, ceea ce joacă un rol semnificativ în procesele de dezvoltare la animale. Un exemplu este pierderea cozii atunci când un mormoloc se transformă într-o broască. Structură: vezicule ovale, membrană în exterior, enzime în interior. Funcții: descompunerea substanțelor organice, distrugerea organelelor moarte, distrugerea celulelor uzate.
Complexul Golgi. Produsele biosintetice care intră în lumenele cavităților și tubilor reticulului endoplasmatic sunt concentrate și transportate în aparatul Golgi. Acest organel măsoară 5-10 μm.

Structura: cavități (bule) înconjurate de membrane. Funcții: acumulare, ambalare, excreție de substanțe organice, formare de lizozomi
Reticulul endoplasmatic. Reticulul endoplasmatic este un sistem pentru sinteza și transportul substanțelor organice în citoplasma unei celule, care este o structură deschisă a cavităților conectate.
De membranele reticulului endoplasmatic sunt atașate un număr mare de ribozomi - cele mai mici organele celulare, în formă de sfere cu un diametru de 20 nm. și constând din ARN și proteine. Sinteza proteinelor are loc pe ribozomi. Apoi proteinele nou sintetizate intră în sistemul de cavități și tubuli, prin care se deplasează în interiorul celulei. Cavități, tubuli, tuburi din membrane, ribozomi de pe suprafața membranelor. Funcții: sinteza substanțelor organice cu ajutorul ribozomilor, transportul substanțelor.
Ribozomi. Ribozomii sunt atașați de membranele reticulului endoplasmatic sau sunt liberi în citoplasmă, sunt localizați în grupuri, iar proteinele sunt sintetizate pe ele. Compoziția proteinelor, ARN ribozomal Funcții: asigură biosinteza proteinelor (asamblarea unei molecule proteice din).
Mitocondriile. Mitocondriile sunt organite energetice. Forma mitocondriilor este diferită; acestea pot fi altele, în formă de tijă, filamentoase, cu un diametru mediu de 1 micron. și 7 µm lungime. Numărul de mitocondrii depinde de activitatea funcțională a celulei și poate ajunge la zeci de mii în mușchii de zbor ai insectelor. Mitocondriile sunt delimitate la exterior de o membrană exterioară, sub care se află o membrană interioară, formând numeroase proiecții - cristae.

În interiorul mitocondriilor se află ARN, ADN și ribozomi. În membranele sale sunt încorporate enzime specifice, cu ajutorul cărora energia nutrienților este transformată în energie ATP în mitocondrii, care este necesară pentru viața celulei și a organismului în ansamblu.
Membrană, matrice, excrescențe - cristae. Funcții: sinteza moleculei ATP, sinteza propriilor proteine, acizi nucleici, carbohidrați, lipide, formarea propriilor ribozomi.
Plastide. Doar în celulele vegetale: leucoplaste, cloroplaste, cromoplaste. Funcții: acumularea de substanțe organice de rezervă, atragerea insectelor polenizatoare, sinteza de ATP și carbohidrați. Cloroplastele au forma unui disc sau o minge cu un diametru de 4-6 microni. Cu o membrană dublă - externă și internă. În interiorul cloroplastei există ADN ribozom și structuri membranare speciale - grana, conectate între ele și cu membrana interioară a cloroplastei. Fiecare cloroplast are aproximativ 50 de boabe, dispuse într-un model de șah pentru a capta mai bine lumina. Membranele Gran conțin clorofilă, datorită căreia energia luminii solare este transformată în energia chimică a ATP. Energia ATP este utilizată în cloroplaste pentru sinteza compușilor organici, în primul rând carbohidrați.
Cromoplastele. Pigmenții roșii și galbeni găsiți în cromoplaste dau diferitelor părți ale plantei culorile lor roșii și galbene. morcovi, fructe de roșii.
Leucoplastele sunt locul acumulării unui nutrient de rezervă – amidonul. Există mai ales multe leucoplaste în celulele tuberculilor de cartofi. La lumină, leucoplastele se pot transforma în cloroplaste (în urma cărora celulele cartofului devin verzi). Toamna, cloroplastele se transformă în cromoplaste, iar frunzele și fructele verzi devin galbene și roșii.
Centrul celular. Este format din doi cilindri, centrioli, situati perpendicular unul pe altul. Functii: suport pentru filete ax

Incluziunile celulare fie apar în citoplasmă, fie dispar în timpul vieții celulei.

Incluziunile dense, granulare conțin nutrienți de rezervă (amidon, proteine, zaharuri, grăsimi) sau produse de deșeuri celulare care nu pot fi încă îndepărtate. Toate plastidele celulelor vegetale au capacitatea de a sintetiza și acumula nutrienți de rezervă. În celulele plantelor, depozitarea nutrienților de rezervă are loc în vacuole.
Boabe, granule, picături Funcții: formațiuni nepermanente care stochează materie organică și energie
Miez. Înveliș nuclear din două membrane, suc nuclear, nucleol. Funcții: stocarea informațiilor ereditare în celulă și reproducerea acesteia, sinteza ARN - informațional, de transport, ribozomal. Membrana nucleară conține spori, prin care are loc schimbul activ de substanțe între nucleu și citoplasmă. Nucleul stochează informații ereditare nu numai despre toate caracteristicile și proprietățile unei celule date, despre procesele care ar trebui să aibă loc în ea (de exemplu, sinteza proteinelor), ci și despre caracteristicile organismului în ansamblu. Informațiile sunt înregistrate în moleculele de ADN, care sunt partea principală a cromozomilor. Nucleul conține un nucleol. Nucleul, datorită prezenței cromozomilor care conțin informații ereditare, funcționează ca un centru care controlează întreaga activitate vitală și dezvoltarea celulei.