Открытые биологические системы. Понятие биологической системы, биологического объекта, гомеостаз
БИОЛОГИЧЕСКАЯ СИСТЕМА - совокупность функционально связанных элементов, образующих целостный биологический объект. Трудность более полного определения Б. с. связана с тем, что это понятие должно отражать основные свойства объективно существующей пространственно-временной структуры, обладающей открытым характером взаимодействия с окружающей средой и имеющей высокую степень специфичности, сложности и организованности.
Один и тот же биол, объект может выступать как в качестве целостной Б. с., так и в качестве ее подсистемы или элемента. Примером биол, объекта, рассматриваемого как Б. с., является организм; на суборганизменном уровне в качестве Б. с. могут рассматриваться клетки, ткани, органы, системы органов (напр., система органов дыхания, пищеварения и т. п.), а на надорганизменном уровне - популяции, экосистемы, биогеоценозы, биосфера. Для последнего типа Б. с. иногда используется специальный термин «биологические макросистемы».
Б. с. обладают высокой устойчивостью к внешним возмущениям; при воздействии факторов среды в них возникают процессы, направленные на уменьшение эффекта этих возмущений. Б. с. сохраняет свою специфичность в изменяющихся условиях окружающей среды и в определенных пределах обеспечивает гомеостаз внутренней среды.
Б. с. являются открытыми системами, т. е. в процессе жизнедеятельности они обмениваются со средой веществом и энергией. Такой обмен является целесообразно регулируемым, управляемым; он направлен на получение свободной энергии из окружающей среды и ее использование для осуществления биол, функций системы. Поэтому для понимания процессов, протекающих в Б. с., необходимо проанализировать работу двух основных составляющих ее частей - энергетической и информационной.
Энергетическая компонента Б. с. связана с процессами получения, накопления, передачи и использования энергии. Эти процессы обеспечивают возможность сохранения структуры, рост, движение и выполнение всех специфических функций Б. с. Вторая часть Б. с. связана с управлением энергетическими процессами и реализует восприятие, хранение, переработку и использование информации. Информационно-управляющие механизмы в Б. с. определяют, какие энергетические процессы и с какой интенсивностью происходят в ней.
Для различных типов Б. с. характерны различные энергетические и информационно-управляющие процессы, но в любой Б. с. обе компоненты тесно связаны между собой и играют существенную роль в поддержании ее существования, совместно обеспечивая сохранение стационарного неравновесного состояния всех подсистем и самой целостной Б. с. Такие процессы наиболее хорошо изучены на организменном и суборганизменном уровне и в несколько меньшей степени - на надорганизменном.
Информационно-управляющие механизмы Б. с. не только обеспечивают стабилизацию и сохранение энергетической части Б. с., но в ряде случаев участвуют в ее образовании. Так, при самовоспроизведении Б. с. организмов генетическое управление формирует как энергетическую компоненту, так и систему информационно-управляющих механизмов физиол, уровня.
Энергетическая компонента биологической системы представляет собой совокупность ее элементов, преобразующих вещество и энергию в процессе жизнедеятельности. Анализ энергетической компоненты Б. с. связан с исследованием структуры и функции органов и тканей, обеспечивающих их энергетический баланс, а на уровне надорганизменных образований - с изучением потоков вещества и энергии в биосообществах и биосфере.
Сложность и специфичность биол, структур и реализующихся в них процессов обусловливает ряд глубоких различий между энергетикой Б. с. и энергетикой неорганического мира. Главной особенностью Б. с. в этом смысле является ее открытый характер. Термодинамика (см.) Б. с. существенно отличается от классической. Фундаментальное для классической термодинамики понятие равновесного состояния заменяется представлением о стационарном неравновесном состоянии системы, а второе начало термодинамики (принцип возрастания энтропии) получает новую формулировку в виде теоремы Пригожина: скорость возрастания энтропии внутри системы стремится к минимальному значению, соответствующему динамическому равновесию.
Изучение энергетической компоненты Б. с. является одной из задач биоэнергетики (см.) - науки, находящейся на стыке биофизики, биохимии и молекулярной биологии.
Информационно-управляющая компонента биологической системы оказывает регулирующее влияние на процессы в энергетической компоненте на всех возможных уровнях организации - от молекулярного до целостного. В целостной Б. с. могут действовать генетические, экологические, эволюционные и физиол, регуляторы, для каждого из которых характерны свои временные интервалы и свои специфические механизмы биол, управления. Так, на уровне целостного организма ведущую и интегрирующую роль в процессах управления и координации функций играют механизмы ц. н. с. В то же время на других уровнях на первый план выступают иные механизмы регуляции (напр., на суборганизменном уровне - специфические Нейро-гуморальные и ауторегуляторные механизмы).
Для осуществления функции управления в Б. с. обязательным является воздействие на контролируемый процесс (или величину, переменную) по меньшей мере двух антагонистических факторов (или процессов), из которых один увеличивает или активизирует данный элемент, а другой - уменьшает или подавляет его. Такими антагонистами могут быть реакции какого-либо метаболического процесса, идущие в прямом и обратном направлении, возбуждающий и тормозной нейроны, мышцы-сгибатели и мышцы-разгибатели и т. д.
Однако, как правило, в Б. с. управляющая функция для нек-рого данного процесса выполняется не просто двумя антагонистическими факторами, а целой группой управляющих механизмов, каждый из которых оказывает регулирующее действие на управляемый процесс в ограниченном диапазоне изменения внешних условий и в нек-рой ограниченной области активности Б. с. Так, напр., тепловой баланс организма поддерживается тем, что вклад в поддержание нормальной температуры вносят два типа регулирующих механизмов - процессы, направленные на выделение тепла (хим. терморегуляция), и процессы, направленные на рассеивание тепла (физ. терморегуляция). В целом взаимодействие этих двух антагонистических факторов приводит к тому, что при любом изменении условий окружающей среды и условий функционирования организма система движется к стационарному состоянию, при к-ром суммарная теплопродукция при всех процессах в организме равна суммарному рассеиванию тепла. Однако каждый из двух главных антагонистических факторов представляет собой результат деятельности целого ряда регуляторных процессов, включающих механизмы дрожи, сосудистых реакций, потоотделения, фосфорилирования, а также поведенческие реакции (поза, мышечная активность).
Наличие множества регуляторных механизмов, дублирующих друг друга и функционирующих параллельно или заменяющих один другой в различных условиях среды, объясняет возможность существования Б. с. в относительно широком диапазоне внешних условий и определяет гибкость и надежность информационно-управляющей системы Б. с.
Изучение кибернетических процессов в информационно-управляющей компоненте Б. с. является задачей биол, и мед. кибернетики, исследующей общие закономерности управления и регуляции в живых системах, находящихся в непрерывном сложном взаимодействии со средой.
Фундаментальным методологическим приемом исследования Б. с. является принцип системной организованности, согласно к-рому любой биол, объект представляет собой Б. с., способную к регулированию как внутренних соотношений между своими подсистемами, так и соотношений целостного объекта со средой. Изучение биол, объектов в таких методологических рамках представляет собой системный анализ биологических систем.
При системном анализе биол, объект представляется в виде Б. с.- множества функционально связанных элементов, реализующих энергетические и информационные процессы в системе. Совокупность существенных связей между этими элементами Б. с. определяет ее структуру. Одним из важнейших свойств сложных систем вообще и Б. с. в частности является иерархичность строения, т. е. последовательное включение систем низкого уровня в системы более высокого уровня. Ряд иерархически соподчиненных систем образует последовательность структурных уровней Б. с.
Рассмотрение Б. с. на различных иерархических уровнях и переход от одного уровня к другому, более сложному, не обязательно связан с усложнением структуры Б. с. и, следовательно, с усложнением ее анализа. Это объясняется тем, что наряду с появлением новых характеристик, не свойственных сумме подсистем предыдущего уровня, ряд признаков этих подсистем оказывается теперь несущественным. Число уровней рассмотрения Б. с. и возможность расчленения Б. с. на элементарные составляющие ограничены.
Низшим уровнем Б. с. является система макромолекул, способных к дискретному воспроизведению, высшим - биосфера, элементами к-рой можно считать биогеоценозы. Между этими уровнями можно выделить несколько промежуточных - клеточный, организменный, популяционный, биогеоценотический.
Можно сказать, что системный анализ представляет собой антитезу известному кибернетическому представлению биол, объектов в виде «черного ящика». Если при исследовании системы как «черного ящика» ученый интересуется только входами (входными процессами или величинами) и выходами системы, а цель исследования состоит в определении закономерностей преобразования входов в выходы без проникновения во внутренние процессы и механизмы системы, то при системном подходе главной целью исследователя является вскрытие внутренней структуры Б. с., понимание реализующихся в этой структуре процессов.
Стандартным методом исследования Б. с. при системном подходе является моделирование ее структуры и закономерностей поведения. При моделировании Б. с. ее энергетическая и информационно-управляющая компоненты рассматриваются совместно как единая целостная система, в к-рой сохранность и функционирование метаболической системы поддерживаются и направляются механизмами регуляции (см. ниже Ауторегуляция в биологической системе). Для математического моделирования управляющих процессов в Б. с. особое значение имеет использование математического и понятийного аппарата кибернетики.
В кибернетической системе наличие регулирования (управления) означает, что ее можно представить в виде двух взаимодействующих блоков - объекта регулирования и регулятора. Регулятор по каналам прямой связи через соответствующее множество эффекторов передает на объект регулирования управляющие воздействия. Информация о состоянии объекта воспринимается рецепторами и по каналам обратной связи (см.) передается в регулятор (схема).
Методологически обоснованным следовало бы считать такое разделение структуры Б. с. на объект и регулятор, при к-ром энергетические и метаболические процессы формируют объект в системе регулирования, а информационно-управляющие процессы связываются с блоком регулятора. Однако представление конкретных Б. с. в виде такой структурной схемы оказывается не всегда возможным, поскольку в сложных живых системах регулятор и объект регулирования структурно и функционально разделяются не всегда, а элементы Б. с. часто совмещают в себе свойства как регулятора, так и объекта регулирования.
В Б. с. имеется два основных типа процессов управления: управление по типу прямой связи (напр., управление по заданной программе - синтез белка на матрице ДНК) и управление по принципу обратной связи. Последний вид управления имеет для Б. с. особое значение.
В простых случаях, когда моделируется влияние какого-либо одного фактора на одну регулируемую переменную, выделяют два типа обратных связей - положительную и отрицательную. Положительная обратная связь увеличивает влияние фактора на выходную переменную, отрицательная - уменьшает. В литературе по моделированию Б. с. получила распространение еще более простая форма описания отрицательной обратной связи - отрицательная обратная связь по отклонению, когда сигнал в цепи обратной связи возникает при отклонении регулируемого сигнала от нек-рого «желаемого» уровня. В сложных Б. с., однако, конкретные регулирующие механизмы и процессы одновременно участвуют в управлении множеством связанных между собой регулируемых процессов, и поэтому для анализа сложных Б. с. в последнее время начинают применяться более сложные модели, разработанные в теории автоматического управления (теория многосвязных систем, теория чувствительности, теория инвариантности).
Понятие «биологическая система» широко используется в таких разделах кибернетики, как кибернетика биологическая и кибернетика медицинская (см.). Стремясь понять природу живого, ученые стремились найти в организме и других биол, объектах то, что можно исследовать изолированно, стремились выделить в них отдельные системы и процессы. Целью же этих разделов кибернетики стало изучение биол, объектов как Б. с. с учетом основных взаимосвязей их структур и элементов. В частности, целый ряд разделов мед. кибернетики опирается на представление об организме как о Б. с. Так, разработка математических методов анализа данных комплексного обследования больного, разработка методов математического моделирования на ЭВМ деятельности его взаимосвязанных функциональных систем, использование автоматизированных машинных методов для оценки состояния больного представляют собой перспективные области, в которых системный анализ может найти непосредственное применение в клинической практике. Системный анализ функций организма как Б. с. позволит сопоставлять и интегрировать показания многочисленных приборов, оценивать общую направленность процессов в организме больного во время больших операций, в послеоперационный и реабилитационный период.
Особое значение работы по исследованию Б. с. на уровне организма приобретают в мед. кибернетике в связи с разработкой новейших методов жизнеобеспечения с применением аппаратов искусственного кровообращения, искусственного дыхания, искусственного сердца. Эффективное использование этих средств приводит к возрастанию требований, предъявляемых к количественным методам в анестезиологии и реаниматологии, поскольку в распоряжении врачей имеется большой выбор средств и методов для восстановления жизненно важных функций больного.
Ауторегуляция в биологической системе
Ауторегуляция в биологической системе - процесс закономерного изменения или поддержания на постоянном уровне определенных регулируемых биол, величин - таких как, напр., кровяное давление, температура тела, митотическая активность клеток тканей, положение конечностей и всего тела в пространстве и т. д. Во многих случаях является очевидным, что эти функции осуществляются соответствующими биол, системами автономно, без внешнего регулирующего воздействия. Поэтому в таких случаях обычно используется более простой термин «регуляция» вместо термина «ауто-регуляция» или других синонимов этого термина.
Различают следующие два основных принципа ауторегуляции: регуляцию по отклонению регулируемой величины и регуляцию по возмущению.
Системы, основанные на первом принципе, способны определять разность между задаваемым и фактическим значением регулируемой величины. Эта разность используется регулятором для выработки регулирующего воздействия на объект, к-рое уменьшает отклонение фактического значения регулируемой величины от заданного значения.
Если задаваемое значение регулируемой величины не изменяется во времени, то ауторегулирующая система представляет собой стабилизатор, т. е. систему, поддерживающую постоянство значения регулируемой величины. Если же задаваемое значение регулируемой величины изменяется во времени по определенному закону, то ауторегулирующая система осуществляет «слежение» за изменением во времени задаваемого значения регулируемой величины и поддерживает минимальную разность между задаваемым значением и текущим (фактическим) значением регулируемой величины.
Отличительной особенностью систем, основанных на принципе регуляции по отклонению регулируемой величины (рис. 1), является наличие у них обратной связи (см.), с помощью к-рой регулятор получает информацию о текущем фактическом значении регулируемой величины. Обратная связь вместе с прямым воздействием регулятора на объект образует замкнутый контур (контур регулирования, контур обратной связи, петлю обратной связи). Поэтому системы, основанные на принципе регуляции по отклонению, часто называют замкнутыми системами регуляции.
Отклонение регулируемой величины от заданного значения, вызванное внешним возмущением, ослабляется в замкнутых системах, т. к. регулятор с помощью обратной связи вырабатывает такое воздействие на объект, к-рое приводит к эффекту, по знаку противоположному первоначальному возмущению. Такая обратная связь называется отрицательной обратной связью.
Представление конкретных Б. с. с автоматическим регулированием в виде единой блок-схемы возможно далеко не всегда. Нередко регулятор и объект регулирования бывают совмещены в одной биол, структуре или функциональной единице.

Рис. 1. Примеры биологических систем автоматического регулирования, основанных на принципе регуляции по отклонению: а - угнетение ключевого фермента E1 конечным продуктом Sn (указано стрелкой с пунктиром), образующимся из исходного субстрата S0 через ряд промежуточных форм S1, S2, ... Sn; Е2, Е3, ... Еn-ферменты, катализирующие промежуточные реакции; стрелкой с штрих-пунктиром показано действие на фермент веществ М, способных активировать или угнетать фермент; б - репрессия синтеза ферментов Е1, Е2, ... Еn конечным продуктом Sn (указано пунктирной стрелкой); Sn угнетает синтез соответствующих информационных РНК (иРНК), препятствуя поступлению необходимой информации для синтеза ферментов, катализирующих промежуточные реакции (Е1. Е2, ... Еn); в - регуляция плотности клеточной популяции в многоклеточном организме. Недифференцированные, способные к делению (стволовые) клетки производят дифференцировавшиеся клетки, оказывающие угнетающее (прямое или косвенное) действие на митотическую активность стволовых клеток. Гормоны, способные усиливать или, напротив, ослаблять угнетение стволовых клеток, определяют задаваемое значение регулируемой величины - плотности популяции дифференцированных клеток.
В частности, угнетение активности ферментов конечными продуктами (угнетение ферментов по принципу обратной связи, ретроингибирование ферментов, аллостерическое угнетение конечными продуктами) состоит в том, что активность фермента, принимающего участие в синтезе данного конечного продукта, уменьшается по мере увеличения концентрации этого продукта (рис. 1, а). Такое угнетение обычно является аллостерическим по своей природе, т. е. угнетающий конечный продукт не имеет стерического (структурного) сходства с субстратом и присоединяется к молекуле фермента в специфическом участке, или центре, отличном от каталитического центра. Такое присоединение влечет за собой конформационную перестройку молекулы фермента и потерю (частичную или полную) активности каталитического центра (см. Конформация). Этот механизм отрицательной обратной связи приводит к стабилизации концентрации конечного продукта: при изменении скорости потребления конечного продукта в широких пределах его концентрация изменяется лишь в относительно узких пределах.
Механизм угнетения ферментов конечными продуктами очень широко распространен. Этот тип регуляции контролирует практически все важнейшие звенья клеточного метаболизма как одноклеточных, так и многоклеточных организмов. Напр., ключевые реакции клеточного метаболизма (гликолиза и окислительного фосфорилирования) угнетаются высокими концентрациями конечного продукта - АТФ. Накопление катионов, сахаров, аминокислот, нуклеотидов, липидов и других жизненно важных веществ в клетке приводит к угнетению ферментов, принимающих участие в активном мембранном транспорте и (или) в биосинтезе этих веществ.
Угнетение, или репрессия, синтеза ферментов конечными продуктами представляет собой значительно более инерционный механизм отрицательной обратной связи по сравнению с механизмом угнетения активности ферментов. Он включается в тех случаях, когда механизм угнетения активности ферментов не в состоянии предотвратить накопление данного конечного продукта в клетке в результате избытка продукта в окружающей среде или резкого снижения скорости его потребления. В таких случаях конечный продукт может вызывать репрессию участка ДНК, несущего наследственную информацию о первичной структуре ферментов, синтезирующих этот продукт (рис. 1, б).
Совместное действие большого числа механизмов отрицательной обратной связи, контролирующих активность и скорость синтеза ферментов, позволяет не только стабилизировать внутриклеточные концентрации многих жизненно важных соединений, но и экономно расходовать исходные субстраты и энергию на синтез этих соединений.
В организации многоклеточных организмов исключительно важную роль играет надклеточный механизм отрицательной обратной связи, контролирующей митотическую активность клеток нормальных тканей. Этот механизм носит название контактного угнетения клеточного деления (син.: угнетение клеточного цикла, контактная регуляция клеточного деления, угнетение клеточного деления, зависящее от плотности, регуляция плотностью и т. д.) и проявляется в том, что митотическая активность нормальных соматических клеток зависит от плотности клеточной популяции, в к-рой они растут. Соматическая клетка при нормальных физиол, условиях обычно не способна делиться и совершать амебоидные движения, если она со всех сторон контактирует с окружающими ее соседними клетками. Устранение этих контактов (полное, а иногда лишь частичное), вызванное удалением или гибелью соседних клеток, вызывает быстрое восстановление способности к активным движениям и делению. Контактное угнетение носит специфический характер и играет важную роль в морфогенетических процессах (см. Морфогенез).
Контактное угнетение, по-видимому, является частным случаем регуляции митотической активности клеток тканей многоклеточного организма. Блок-схема такой регуляции, в к-рой главную роль играет угнетающее действие дифференцировавшихся клеток (прямое или косвенное) на митотическую активность недифференцировавшихся (стволовых) клеток, показана на рисунке 1, в. Типичным примером такой схемы ауторегуляции является регуляция численности кровяных клеток.
Системы, основанные на принципах регуляции по возмущению, используют само возмущение для выработки нек-рого компенсирующего воздействия на объект для того, чтобы предотвратить изменение регулируемой величины. Поскольку фактическое значение регулируемой величины в таких системах не оказывает воздействия на регулятор, то в них отсутствует замкнутый контур регулирования, характерный для систем с отрицательной обратной связью (регулирование по отклонению). Это обстоятельство делает практически невозможной полную компенсацию возмущения, что приводит к изменению регулируемой величины в сторону, противоположную той, в к-рую изменяется эта величина при отсутствии регулятора.
При некоторых условиях компенсирующее воздействие может вызывать даже большее отклонение регулируемой величины, чем нескомпенсированное возмущение.
![]()
Наиболее широко изученными и наиболее распространенными механизмами ауторегуляции компенсационного типа на молекулярном уровне являются механизмы, с помощью которых исходные субстраты оказывают активирующее действие на ферменты, принимающие участие или в утилизации этих субстратов (субстратная активация, опережающая активация, активация предшественниками субстрата), или на скорость синтеза таких ферментов (индукция ферментов, индукция синтеза ферментов, субстратная индукция ферментов).
Действие механизма активации фермента предшественником субстрата можно пояснить с помощью схемы, представленной на рисунке 2, а.
На схеме показана последовательность реакций, в к-рой исходный субстрат S0 превращается через ряд промежуточных форм (S1, S2... и т. д.) в соединение Sn, утилизируемое различными реакциями. Это соединение служит субстратом для фермента Еk, чувствительного к активирующему действию исходного субстрата S0 (показано пунктирной линией). Изменение скорости образования исходного субстрата V0 действует как внешнее возмущение на концентрацию Sn: при увеличении V0 эта концентрация также увеличивается. Однако активация фермента Еk посредством S0 позволяет скомпенсировать усиленное образование Sn из S0 за счет увеличения оттока Sn в реакцию, катализируемую этим ферментом. При неизменной активности ферментов, утилизирующих Sn, и при определенных соотношениях параметров системы реакций возможна практически полная компенсация возмущения, вызванного изменением V0: концентрация Sn может оставаться почти неизменной при изменении V0 в широких пределах.
Простейшей формой регуляции активности ферментов компенсационного типа является аллостерическая активация ферментов субстратами.
Молекулы субстратов-активаторов присоединяются к регуляторным центрам на молекуле фермента, что вызывает такое изменение его конформации, к-рое способствует увеличению каталитической активности активного центра.
Индукция синтеза ферментов субстратами (рис. 2, б) представляет собой инерционный аналог механизма субстратной активации ферментов.
Сущность явления состоит в том, что субстраты могут вызвать дерепрессию синтеза ферментов, участвующих в утилизации этих субстратов. Индукция синтеза ферментов является одним из универсальных механизмов адаптации клеток к меняющимся условиям внешней среды. Нередко в этот процесс вовлекается естественный отбор, который приводит в конечном счете к появлению клеток, способных синтезировать новые ферменты. Типичным примером такого сорта ауторегуляции, использующей естественный отбор, может служить адаптация бактерий к антибиотикам и насекомых к инсектицидам. В частности, у насекомых, подвергавшихся в течение многих поколений действию инсектицида ДДТ, выработалась способность синтезировать новый фермент - ДДТ-дехлоргидразу, инактивирующий этот инсектицид.
Примером ауторегуляции компенсационного типа на физиол, уровне может служить регуляция уровня глюкозы в крови поджелудочной железой (рис. 2, в). Повышение поступления глюкозы из кишечника стимулирует образование инсулина поджелудочной железой, который стимулирует депонирование глюкозы в форме гликогена и жиров в печени, мышцах и жировой ткани. Этот усиленный отток глюкозы в депонирующие ткани компенсирует усиленный приток глюкозы из кишечника. Подобные компенсационные механизмы прослеживаются в регуляции секреторной активности всех эндокринных желез и в иммунных реакциях организма. Компенсационные механизмы ауторегуляции являются основой очень многих поведенческих реакций, контролируемых ц. н. с., и проявляются обычно в форме рефлекторного устранения раздражителя.

Рис. 3. Комбинированные системы автоматического регулирования: а - блок-схема комбинированной системы автоматического регулирования (регулятор I реагирует на возмущение, регулятор II - на отклонение от заданного значения); б - простая биохимическая система ауторегуляции комбинированного действия. Угнетение фермента Et продуктом S представляет собой механизм отрицательной обратной связи, а активация фермента Е2 субстратом S - компенсационный механизм. Действие обоих механизмов направлено на стабилизацию уровня вещества S, используемого потребителями; в - комбинированная многоконтурная система регулирования уровня глюкозы в крови. На схеме представлены два контура: отрицательной обратной связи (пунктир) и компенсирующей связи (штрих-пунктир).
Комбинированные системы ауторегуляции. В большинстве своем Б. с. представляют собой комбинированные системы ауторегуляции. Такие системы имеют в своей структуре как механизмы отрицательной обратной связи, так и механизмы компенсации возмущения (рис. 3, а). С точки зрения теории автоматического регулирования такие комбинированные системы способны обеспечить почти идеальную изоляцию регулируемого объекта от внешних возмущений. На рисунке 3, б приведена схема типичной комбинированной биохим. системы ауторегуляции, содержащей отрицательную обратную связь (угнетение фермента его продуктом S) и компенсационную связь (активацию фермента Е2 его субстратом S). Действие обоих механизмов направлено на стабилизацию уровня вещества S, используемого потребителем.
Все важнейшие промежуточны© соединения клеточного метаболизма контролируются подобными комбинациями регулирующих связей. Напр., гексозомонофосфаты угнетают свое образование в реакциях, катализируемых ферментами гексокиназой, глюкокиназой, фруктокиназой и гликогенфосфорилазой, и активируют свою утилизацию в реакциях, катализируемых ферментами глюкозо-6-фосфатдегидрогеназой, фосфофруктокиназой и гликогенсинтетазой. Аналогичным образом регулируется в клетках уровень и других соединений (напр., АМФ, ацетил-КоА, многих интермедиатов цикла Кребса, большинства аминокислот).
Принцип комбинированной ауто-регуляции широко используется и на более высоких уровнях биол, организации. На рисунке 3, в приведена блок-схема нейрогуморальной регуляции уровня глюкозы в крови. На схеме представлены три ауторегулирующих механизма. Два из них представляют собой отрицательные обратные связи, включающиеся при понижении уровня глюкозы. Оба этих механизма приводят к активации образования глюкозы в печени из гликогена печени и лактата, аминокислот и глицеридов, транспортируемых в печень из мышц и жировой ткани. Третий, компенсирующий механизм включается при увеличении уровня глюкозы в крови. Действие этого механизма сводится к активации депонирования глюкозы в различных тканях. Кроме указанных на схеме механизмов, существует еще ряд нейрогуморальных механизмов, контролирующих уровень глюкозы. Среди таких механизмов большую роль в регуляции уровня глюкозы играет механизм повышения уровня глюкозы, замыкающийся через цепь: гипоталамус -> гипофиз -> АКТГ -> мозговой слой надпочечников -> адреналин -> активация фосфорилазы печени -> увеличение глюкозы крови по принципу отрицательной обратной связи.
Все Б. с. ауторегулирования обладают определенной инерционностью, из-за к-рой быстрое (напр., скачкообразное) возмущение регулируемого объекта не может быть мгновенно скомпенсировано регулятором, и поэтому регулируемая величина успевает отклониться (иногда довольно сильно) от заданного значения прежде, чем регулятор сработает соответствующим образом. Процесс достижения регулируемой величиной заданного стационарного значения, вызванный однократным возмущением регулируемого объекта, называется переходным процессом.
Ауторегуляция, порождающая автономные движения в биологических системах. Фундаментальным свойством живых систем является их способность к различным видам автономных (не зависящих от внешних возмущений) изменений биол, величин во времени и пространстве. Все виды эндогенных биол, ритмов, все формы механического движения, генерация и проведение нервных импульсов, переработка информации в нервной системе и т. д.- все это прямо или косвенно является результатом функционирования особых механизмов ауторегуляции - генераторов автономных движений.
Самодвижение любой системы вызывается и поддерживается неустойчивостью равновесного или стационарного состояния системы. Поэтому характерной особенностью генераторов автономных движений является наличие у них специального механизма, обеспечивающего такую неустойчивость,- так наз. положительной обратной связи. Положительная обратная связь вырабатывает такое воздействие на регулируемый объект, к-рое совпадает по знаку с первоначальным отклонением регулируемой величины от стационарного значения, в соответствии с чем состояние систем с положительной обратной связью при определенных условиях оказывается неустойчивым. Сколь угодно малое возмущение регулируемой величины, вызванное внешним воздействием или неизбежными флюктуациями (т. е. самопроизвольными колебаниями) в самой системе, приводит к прогрессивно нарастающему отклонению регулируемой величины от исходного значения. Отклонение продолжается до тех пор, пока система не достигнет нового, уже устойчивого стационарного состояния. Этим новым стационарным состоянием может быть либо состояние покоя, в к-ром переменные не изменяются во времени, либо автоколебательное состояние, в к-ром переменные колеблются во времени с постоянной амплитудой.
Типичным механизмом положительной обратной связи на молекулярном уровне является аутокатализ (см.). Очень многие звенья клеточного метаболизма носят аутокаталитический характер. Напр., для синтеза АТФ необходимо, чтобы АТФ уже присутствовал в каталитических количествах в клетках. Для «запуска» фосфорилирования АДФ гликолитической системой необходимо наличие АТФ, который служит кофактором первых двух стадий. Положительная обратная связь контролирует активность таких ферментов, как фосфорилаза - ключевой фермент гликогенолиза, глутаматдегидрогенеза - ключевой фермент азотистого обмена.
Размножение вирусов, одноклеточных и многоклеточных организмов при отсутствии сдерживающих размножение факторов также носит аутокаталитический характер.

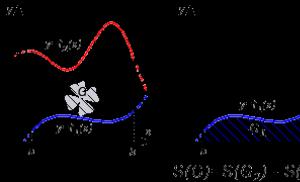
Рис 4. Множественность стационарных состояний в биологической системе с положительной обратной связью: а - система с положительной обратной связью, состоящая из источника вещества P и потребителя этого вещества. Вещество P образуется источником со скоростью V (p) и и утилизируется потребителем со скоростью V (p) п. Штрих-пунктиром показана положительная обратная связь - активирующая действие P на источник; б - графическое изображение взаимодействия источника и потребителя. Поскольку скорость образования продукта описывается S-образной кривой, взаимодействие источника и потребителя порождает три альтернативных стационарных состояния, представленных на графике точками пересечения кривых, описывающих зависимость скоростей и от концентрации P (жирные кривые V (p) и и V (p) п. Каждому стационарному состоянию (точки 1, 2 и 3) соответствует свое значение концентрации P (концентрации [P]1, [Р]2 и [Р]3) и свое значение стационарной скорости V = V (p) и = V (p) п (значения V1, V2 и V3). Пунктиром показана зависимость скорости V (p) и от концентрации P при отсутствии положительной обратной связи.
Множественность альтернативных стационарных состояний в биологической системе. Простейшей формой автономного движения является переключение системы из одного стационарного состояния в другое. Подобное переключение может быть изучено на примере системы с положительной обратной связью, показанной на рисунке 4, а. Эта система состоит из источника и потребителя какого-либо продукта (Р). Образование P регулируется положительной обратной связью. Скорость образования P источником увеличивается с увеличением концентрации Р. Если скорость утилизации P потребителем растет с увеличением концентрации P (рис. 4, б), то в системе источник - потребитель могут существовать три альтернативных стационарных состояния, характеризующиеся тремя различными значениями концентрации P и тремя различными значениями скоростей и V (p) п. На рисунке стационарные состояния представлены точками пересечения графиков, изображающих зависимость скоростей V (p) и и V (p) п от концентрации P (точки 1, 2 и 3). В каждой такой точке выполняется условие:

т. е. концентрация P не меняется со временем t.

При малых отклонениях концентрации P в обе стороны от стационарных значений [P]1 и [Р]3 разность скоростей - V (p) и - V (p) п компенсирует эти отклонения так, что концентрация P снова принимает прежнее стационарное значение. Однако при сколь угодно малом отклонении концентрации P от стационарного значения [Р]2 разность скоростей V (p) и - V (p) п за счет действия положительной обратной связи приводит к дальнейшему отклонению концентрации P от значения [Р]2 в сторону, совпадающую с первоначальным отклонением. Так, при малом увеличении [Р] по сравнению с [Р]2 разность V (p) и - V (p) п оказывается положительной, и [Р] начинает увеличиваться со временем - в системе начинается автономное движение, приводящее к переключению системы из неустойчивого стационарного состояния (рис. 5,2) в устойчивое состояние (рис. 5,7). При малом отклонении [Р] в сторону уменьшения по сравнению со стационарным значением [Р]2 разность скоростей V (p) и - V (p) п <0, и это приводит к дальнейшему уменьшению концентрации P до тех пор, пока она не примет нового устойчивого значения [Р]1 (кривая 2).

Рис. 6. Критические явления и гистерезис в системе с положительной обратной связью: а - система с положительной обратной связью, в которой скорость образования продукта P источником V (p) и зависит от концентрации субстрата S V (p) п - скорость утилизации P потребителем; б - взаимодействие источника и потребителя при нескольких фиксированных значениях концентрации S (значения концентрации S указаны на графике в условных единицах). Точки пересечения кривых V (p) и и кривой V (p) п представляют со бой стационарные состояния системы. Число стационарных состояний изменяется с изменением концентрации S; в - зависимость стационарной скорости V = V (p) и = V (p) п =от концентрации субстрата S. Стрелками показаны направления автономных движений при достижении концентрации S одного из двух критических значений - при [S] = 4 переход из точки а в точку в, при [S] = 2 переход из точки с в точку d.
Изменения параметров, определяющих скорость источника V (p) и и скорость потребителя V (p) п, могут смещать стационарные состояния - изменять стационарные значения скоростей и концентрации Р. При достижении некоторых критических (пороговых) значений могут исчезать и появляться новые стационарные состояния, а система переключается из одного стационарного состояния в другое. Если скорость источника V (p) и зависит от концентрации субстрата S, из к-рого образуется продукт P (рис. 6, а), то изменение концентрации S меняет взаимное расположение стационарных точек (рис. 6, б), где представлено семейство кривых V (p) и построенных при пяти фиксированных значениях концентрации субстрата S, и кривая V (p) п. При малой концентрации S, т. е. 0<= [S] < 2, в системе существует только одно стационарное состояние. Это состояние устойчиво, как и состояние 1 на рисунке 4, б, и характеризуется низкими стационарными значениями концентрации P и скоростей V (p) и и V (p) п. При высокой концентрации S - ([S] >4) - в системе также существует одно стационарное состояние, к-рое, как и состояние 3 (рис. 4, б), устойчиво, но характеризуется высокими стационарными значениями переменных. При средних значениях концентрации S - (2 < [S] < 4) - в системе существуют три стационарных состояния, подобные состояниям 1, 2 и 3 (рис. 4, б). Неустойчивое промежуточное стационарное состояние [средняя точка пересечения кривой V (p) п построенной для [S] = 3, с кривой V (p) и (рис. 6, б)] с увеличением концентрации сближается с верхним устойчивым стационарным состоянием, а при уменьшении концентрации S - с нижним устойчивым стационарным состоянием. Существует два критических значения концентрации S ([S] - 2 и [S] = 4), при которых неустойчивая стационарная точка сливается с устойчивой верхней (при [S] = 2) или нижней (при [S] = 4) стационарной точкой, в результате чего устойчивость этих точек теряется, и система совершает автономное движение - переключается в одну из устойчивых стационарных точек. На рисунке 6,в показана зависимость стационарной скорости V = V (p) и = V (p) п от концентрации субстрата. Эта зависимость носит гистерезисный характер: прямой и обратный переходы системы из одного устойчивого стационарного состояния в другое (показаны на рисунке стрелками) совершаются при различных критических значениях концентрации S и разными путями. Благодаря этому свойству система оказывается неэквифинальной: в зависимости от начальных условий она может оказаться в одном из двух устойчивых альтернативных стационарных состояний. Т. о., система оказывается способной неопределенно долго хранить след (помнить) о таких изменениях в среде, которые способны приводить к смене ее стационарных состояний. Емкость памяти рассмотренной системы минимальна и равна 1 биту. Однако большое число подобных неэквифинальных систем, функционирующих в большой Б. с., может служить основой памяти большой емкости. Системы, имеющие два устойчивых стационарных состояния, часто называют триггерами с двумя устойчивыми состояниями (иногда просто триггерами) или бистабильными системами.
Автоколебания . Если в системе (рис. 6, а) весь субстрат превращается в продукт Р, то скорость потребления продукта (V (s) п) равняется скорости его образования (V (p) п), и поэтому зависимость V (s) п от [S] имеет точно такой же вид (рис. 6, б), как и зависимость V от [S] на рисунке 6, в. Гистерезисная зависимость от концентрации S может быть причиной одного из важнейших видов автономных движений - автоколебаний (автономных колебаний).

Рис. 7. Автоколебания в системе с положительной обратной связью: а - система с положительной обратной связью, состоящая из источника субстрата S, потребителя субстрата, служащего источником продукта Р, и потребителя продукта Р. Продукт P активирует свое собственное образование. Скорости образования S и P обозначены через V (s) и и V (p) и, а скорости утилизации - через V (s) п и V (p) п; б - взаимодействие источника и потребителя субстрата S. S-образная кривая представляет собой зависимость скорости V (s) п от концентрации S, а прямая V (s) и - зависимость скорости от концентрации S. Пересечение кривой V (s) п с прямой V (s) и (точка о) представляет собой стационарную точку системы. Замкнутый контур b,с,о,a, d представляет собой предельный цикл, по которому система совершает автономное циклическое движение; в - автоколебания скорости V (s) п во времени t. Буквами отмечены значения скорости, соответствующие точкам предельного цикла, показанного на рисунке 7, б слева.
Автоколебания возникают в том случае, если субстрат образуется нек-рым источником (рис. 7, а) с такой скоростью (V (s) и), что стационарное состояние системы, определяемое двумя условиями

оказывается неустойчивым. При гистерезисном характере зависимости V (s) п от концентрации S стационарное состояние (точка О на рис. 7, б) неустойчиво, если значения скорости источника V (s) и совпадают с неустойчивыми значениями скорости V (s) п. Если стационарное состояние неустойчиво, то при малейшем отклонении концентраций S или P от стационарных значений система начинает автономное движение к одной из устойчивых ветвей кривой V (s) п (движение О->b" или О->d" на рис. 7, б). В момент достижения устойчивой ветви (точка b" или точка d") в системе устанавливается квазистационарное состояние, в к-ром удовлетворяется условие (3), но не условие (2). Это квазистационарное состояние характеризуется медленным дрейфом системы вдоль устойчивой ветви кривой V (s) п по направлению к одному из экстремумов этой кривой (движение b" -> с или d" -> а). В момент достижения экстремальной точки (с или а) система «срывается» в быстрое движение (с -> d или а -> b) по направлению к другой устойчивой ветви. После достижения устойчивой ветви движение снова резко замедляется и опять начинается медленный дрейф (d -> а или b -> с). Т. о., гистерезис зависимости V (s) п от концентрации S и неустойчивость стационарного состояния приводят к автономному движению системы по замкнутому циклу (по циклу а -> b -> с -> d ->а). Такой цикл называют предельным циклом системы. При движении системы по предельному циклу все ее переменные периодически изменяются. На рисунке 7, в показаны автономные колебания (автоколебания) скорости V (s) п во времени t.
Кроме положительной обратной связи, имеется большое разнообразие механизмов ауторегуляции, способных вызывать неустойчивость стационарных состояний Б. с. и тем самым служить основой для генерации различных форм автономных Движений. В частности, системы с отрицательной обратной связью при определенных условиях также могут терять устойчивость и быть источником автоколебаний. Основной причиной потери устойчивости такими системами является чрезмерная инерционность механизма обратной связи и слишком большая чувствительность регулятора к сигналу обратной связи, приводящие к тому, что регулирующее воздействие большой амплитуды вырабатывается регулятором с большим запаздыванием, что вызывает сильное перерегулирование - отклонение регулируемой величины в сторону, противоположную первоначальному возмущению. Это вторичное отклонение регулируемой величины приводит в свою очередь к выработке нового регулирующего воздействия, к-рое из-за запаздывания вновь вызывает перерегулирование, но уже противоположного знака, и т. д. Такое «рыскание» системы ауторегуляции с отрицательной обратной связью при определенных условиях может иметь автоколебательный характер.
Автоколебательные процессы наблюдаются на всех уровнях организации живой материи. Колебания концентраций реагирующих веществ могут возникать в отдельных ферментативных реакциях, полиферментных системах, мембранном транспорте, мышечном сокращении, митотической активности клеток различных тканей, электрической активности коры головного мозга и т. д. Автоколебания, генерируемые различными Б. с., служат основой временной организации (т. е. определенного порядка функционирования во времени) Б. с. Автоколебания играют очень важную роль в процессах кодирования, передачи и преобразования информации в нервной системе. Напр., большинство рецепторных клеток является автоколебательными системами, кодирующими информацию об измеряемых параметрах среды (интенсивность света, температура, давление, концентрация различных веществ и т. д.) в форме автоколебаний мембранного потенциала соответствующей амплитуды и частоты.

Рис. 8. Генерация импульса стандартной формы в системе с положительной обратной связью: а - зависимость скорости потребления субстрата V (s) и и скорости образования субстрата источником V (s) и от концентрации субстрата S; б - изменение скорости потребления субстрата V (s) п во время t0, вызванное кратковременным сверхпороговым увеличением скорости источника V (s) и в момент времени t0. Продолжительность возбуждающего действия увеличения V (s) и показана отрезком под графиком. Штрих-пунктиром показан критический уровень V (s) и, превышение которого приводит к генерации системой импульса стандартной формы. Жирной пунктирной кривой показано изменение V (s) п, вызванное кратковременным подпороговым увеличением V (s) и, продолжительность которого указана короткой жирной чертой под графиком. Стационарное значение V (s) п показано пунктирной прямой.
Генерация одиночного импульса стандартной формы. Если стационарная точка системы, показанной на рисунке 7, а, расположена на устойчивой ветви кривой V (s) п вблизи одного из экстремумов, как это показано на рисунке 8, а, то такое состояние характеризуется относительной устойчивостью. При малых возмущениях система возвращается в исходное состояние. Однако при больших, сверхпороговых возмущениях система временно теряет устойчивость и совершает однократное автономное циклическое движение (а -> b -> с -> d -> о, рис. 8, а), завершающееся возвратом в исходное стационарное состояние. В результате такого цикла система генерирует однократный импульс (рис. 8, б), амплитуда и длительность к-рого практически не зависят от амплитуды и длительности сверхпорогового возмущения. Такое поведение типа «все или ничего» (см. «Все или ничего», закон) характерно для многих Б. с., в частности для биологических мембран, способных возбуждаться под действием сверхпорогового стимула.
Переключение из одного стационарного состояния в другое, генерация однократных импульсов стандартной формы и генерация автоколебаний являются элементарными формами автономных движений. Взаимодействие элементарных движений друг с другом в Б. с., содержащих большое число различных генераторов автономных движений, порождает сложные формы движений. Примером Б. с., генерирующей такие сложные формы движений, является нервная система.
Патология как нарушение нормального функционирования систем ауторегуляции
Все виды Б. с. ауторегуляции способны выполнять свои нормальные функции гомеостаза или генерации автономных движений только в определенных диапазонах условий. Возмущения окружающей среды и процессы внутреннего развития (в частности, старения) организма могут вызывать изменения внутренней среды, не компенсируемые гомеостатическими механизмами. Под влиянием таких некомпенсируемых изменений происходят смещения стационарных состояний различных систем ауторегуляции. Эти смещения, достигая критических границ, вызывают потерю устойчивости стационарных состояний и переключение систем в новые стационарные состояния и генерацию ими новых типов автономных движений. Новые стационарные состояния и автономные движения могут быть несовместимыми с нормальным функционированием организма, и тогда в организме развивается патологическое состояние.
Надежность систем ауторегуляции. Б. с. ауто-регуляции характеризуются особыми формами организации, благодаря к-рым достигается исключительно высокая надежность функционирования таких систем (см. Надежность). Особенностью надежности Б. с. по сравнению с техническими является высокая надежность всей системы в целом при относительно низкой надежности входящих в нее элементов (клетки, органеллы и т.д.). Важнейшими чертами этой организации являются: 1) множественность (дублирование) регулирующих связей, контролирующих одну и ту же функцию; 2) множественность и относительная автономность параллельно функционирующих элементов; 3) многократное резервирование, депонирование запасных питательных веществ и источников энергии; 4) замещение или восстановление элементов и связей, вышедших из строя; 5) способность к самоочищению от чужеродных элементов; 6) защита от перегрузок и способность адаптироваться к частым перегрузкам.
Библиография: Аптeр М. Кибернетика и развитие, пер. с англ., М., 1970; Б Ηχο в с к и й М. JI. и Вишневский А. А. Кибернетические системы в медицине, М., 1971, библиогр.; Воронов А. А. Основы теории автоматического управления, ч. 3, М.- Л., 1970, библиогр.; Новосельцев В. Н. Гомеостаз систем управления, Автоматика и телемеханика, № 12, с. 118, 1973, библиогр.; УотерменТ. Теория систем и биология, в кн.: Теория систем и биология, пер. с англ., под ред. В. И. Кринского, с. 7, М., 1971, библиогр.; Шумаков В. И. и др. Моделирование физиологических систем организма, М., 1971.
Ауторегуляция в Б. с. - Клегг П. и Клегг А. Гормоны, клетки, организм, пер. с англ., М., 1971, библиогр.; Колебательные процессы в биологических и химических системах, под ред. Г. М. Франка, М., 1967, библиогр.; М о-н о Ж. и Ш а к о б Ф. Общие выводы, телеономические механизмы в процессах клеточного обмена, в кн.: Регуляторн. механизмы клетки, пер. с англ., под ред. И. Б. Збарского, с. 477, М., 1964, библиогр.; Сельков E. Е. Исследование механизма гликолитических колебаний, в кн.: Математ. модели биол, систем, под ред. Г. М. Франка, с. 5, М., 1971, библиогр.; T а с а к и И. Нервное возбуждение, пер. с англ., М., 1971, библиогр.; Тринкаус Д. От клеток к органам, пер. с англ., М., 1972, библиогр.; X а-руна И. и СпигельманС. Авто-каталитический синтез вирусной РНК, в кн.: Ферменты и синтез биополимеров, пер. с англ., под ред. Я. М. Варшавского, с. 90, М., 1967; Stadtman E. R. Allosteric regulation of enzyme activity, Advanc. Enzymol., v. 28, p. 41, 1966, bibliogr.; он же, Mechanism of enzyme regulation in metabolism, в кн.: Enzymes, ed. by P. D. Boyer, v. 1, p. 397, N. Y., 1970.
В. H. Новосельцев; E. E. Сельков (биофиз.).
Концепция системной многоуровневой организации жизни — одна из ключевых в современном естествознании. Все биологические объекты, согласно ей, объединяются на основе некоторых признаков и тесных взаимоотношений и выстраиваются в определенном иерархическом порядке. Схожие принципы универсальны для всей природы в целом. Знакомство с тем, что такое биологические системы, лучше начать с определения ключевого понятия.
Всеобъемлющая теория
Основы концепции были заложены в середине прошлого столетия Людвигом фон Берталанфи. Именно он разработал общую теорию систем. Она охватывает все и общества. Теория выделяет биологические, социальные, космические, физические, экономические и прочие системы, объединяющиеся в три крупные категории: микромир, макромир и мегамир. К первому относятся элементарные частицы и атомы, ко второму — все, от молекул до океанов и материков, к третьему — космические объекты. Макромир включает и живые системы.
Основное понятие
Система — объединение элементов, базирующееся на определенных взаимоотношениях, подчиненных некоторым законам. Организация подобной структуры, как правило, состоит из нескольких упорядоченных уровней. При этом каждый элемент может одновременно быть и системой менее высокого порядка. Важное свойство подобной организации: целое качественно отлично от суммы всех своих составляющих. Система - не просто набор характеристик элементов, она отличается неким новым качеством.
Все объекты живого мира представляют собой подобные структуры. Причем качеством, возникающим в результате объединения нескольких элементов, становится новое проявление жизни.

Открытые
Понимание того, что такое биологические системы, требует описания еще одного свойства подобных структур. Это взаимодействие с окружающей средой. В биологических систем может быть как замкнутой, так и открытой. На практике ученым не известно ни одной полностью закрытой структуры. Любая живая система постоянно взаимодействует с окружающей средой через некоторую полупроницаемую пограничную оболочку. У клеток — это билипидная мембрана, у космической станции — обшивка. объединяются посредством законодательных актов или определенных взаимоотношений людей.
Получается, ответ на вопрос «Что такое биологические системы?» можно сформулировать следующим образом: это совокупность постоянно взаимодействующих живых элементов, выстроенная в определенном иерархическом порядке и открытая в той или иной степени для обмена с окружающей средой.
Признаки
Все отличительные характеристики рассматриваемых структур — это одновременно и критерии отличия живой природы от неживой. Назовем признаки биологических систем с их краткой характеристикой:
- Единый химический состав. Все природные объекты построены из одних и тех же молекул. Однако живая материя в качестве основных элементов включает углерод, азот, кислород и водород.
- Обмен веществ со средой. Это уже описанное свойство открытости системы. Одно из его проявлений — энергозависимость подобных структур.
- Самовоспроизведение (размножение).
- Наследственность — свойство передавать особенности строения и функционирования из поколения в поколение.
- Изменчивость — свойство приобретать в течение жизни новые характеристики и навыки.
- Рост и развитие. Представляют собой направленное необратимое изменение. Выделяют индивидуальное и историческое развитие живых систем, называемые онтогенезом и филогенезом соответственно.
- Раздражимость (рефлексы, таксисы) — свойство реагировать на стимулы и изменения окружающей среды.
- Дискретность. Любая живая система состоит из отдельных, но взаимодействующих элементов, образующих иерархическую структуру.
- Саморегуляция. Существуют внутренние механизмы поддержания гомеостаза, способствующие выживаемости системы. Саморегуляция основана на принципе отрицательной обратной связи.
- Ритмичность. Усиление и ослабление различных процессов через равные промежутки времени.
Уровни организации биологических систем

Все описанные свойства сохраняются на любой ступени иерархической структуры. Основные уровни организации биологических систем выделяются достаточно условно, поскольку любой из них легко разделить на несколько составляющих. В общем случае говорят о четырех ступенях этой иерархии:
- молекулярно-генетический уровень;
- онтогенетический уровень;
- популяционно-видовой уровень;
- биогеоценотический уровень.
Остановимся на них подробнее.
Молекулярно-генетический уровень
Такие макромолекулы, как белки, липиды, углеводы и нуклеиновые кислоты, представляют собой структурные элементы организмов, но сами по себе не являются носителями полноценной жизни.

Каждый из названных элементов выполняет свои функции. Углеводы — источник энергии. Липиды входят в состав клеток. Также они являются поставщиком энергии. Белки выполняют большую часть жизненных функций. Они состоят из двадцати разновидностей аминокислот, которые могут чередоваться в произвольном порядке. В результате существует огромное количество белков, способных справляться с самой разной работой. Нуклеиновые кислоты, ДНК и РНК, — основа наследственности.
Макромолекулы объединяются в комплексы, образуя органоиды клетки: рибосомы, митохондрии, миофибриллы и так далее. Все они отвечают за отдельные биологические проявления, однако не достигают того уровня сложности, который можно назвать жизнью.
Следующая ступень
Какие биологические системы составляют онтогенетический уровень? Это все организмы, начиная от одноклеточных и заканчивая млекопитающими и человеком, а также органы, ткани и клетки тела. Все названные элементы могут рассматриваться как отдельные уровни организации биологических систем, однако для удобства и в силу общих закономерностей они объединены.

Клетка — элементарная структурная единица строения организмов. Она же представляет собой тот уровень сложности биологической системы, на котором впервые возникает жизнь как явление. Как уже было сказано, более простые структуры обеспечивают лишь отдельные функции. Клетке же присущи все свойства биологических систем.
Ткани и органы — промежуточные подуровни онтогенетической ступени. За ними идет многоклеточный организм. Он характеризуется способностью к самостоятельному существованию, развитию и размножению. Это свойство отличает особь и клетку от органов и тканей.
Популяция и вид
От ступени к ступени происходит усложнение биологических систем. На следующем уровне располагаются виды и популяции. Первые представляют собой совокупность особей, характеризующихся наследственным сходством по целому ряду параметров: морфология, физиология, генетика, географическое размещение. И самое главное: организмы, составляющие вид, способны свободно скрещиваться и оставлять плодовитое потомство.
Группа особей занимает определенную территорию, которую называют ареалом обитания. Достаточно часто он бывает разорван различными географическими препятствиями. В результате вид распадается на несколько относительно изолированных популяций. Естественно, что условия отъединения от остального вида способствуют накоплению определенного генетического материала. При сильном расхождении признаков популяций появляются новые виды.
Экосистемы

В иерархической лестнице за популяциями и видами следует сообщество, биогеоценоз и биосфера. Первое представляет собой совокупность популяций разных видов, размещающихся на одной территории. Выделяют растительные, животные и микробные сообщества. Их совокупность в пределах одного ареала будет называться биоценозом. Эти уровни биологических систем характеризуются тесной взаимосвязью всех особей.
Условия, в которых существуют организмы, постоянно влияют на них. Всю совокупность подобных данного ареала принято называть биотопом. Среда и сообщества организмов пребывают в постоянном взаимодействии, происходит круговорот вещества и энергии. Поэтому биотоп и биоценоз объединяют в биогеоценоз, или экосистему. Этот уровень также характеризуется всеми особенностями живого: он постоянно контактирует со средой, управление в нем происходит по принципу саморегуляции, процессы подчиняются определенным циклам.
На высшей ступени иерархии размещает биосфера Земли — оболочка, населенная живыми существами. Огромное влияние на нее оказывает деятельность человека, что все чаще приводит к возникновению экологических катастроф.
Что такое По сути, это все то живое, что нас окружает. Человек отличается от других элементов в биосфере возможностью осознавать, а значит, перенаправлять и изменять свою деятельность. Пока эта способность Homo sapiens работает против природы. Однако именно благодаря ей у нас есть шанс все исправить.
Понятие открытой системы ввел Л. фон Берталанфи. Основные отличительные черты открытых систем - способность обмениваться со средой массой, энергией и информацией. К ним безусловно относятся биологические системы.
Одна из наиболее полных и интересных классификаций по уровням сложности предложена К. Боулдингом. Выделенные в ней уровни приведены в табл. 1.
|
О казино вулкан отзывы пишет достаточно много игроков казино вулкан автоматы , вступайте. Тип системы |
Уровень сложности | |
|
Живые системы |
Открытые системы с самосохраняемой структурой (первая ступень, на которой возможно разделение на живое и неживое) |
Клетки, гомеостат |
|
Живые организмы с низкой способностью воспринимать информацию |
Растения |
|
|
Живые организмы с более развитой способностью воспринимать информацию, но не обладающие самосознанием |
Животные |
|
|
Системы, характеризующиеся самосознанием, мышлением и нетривиальным поведением | ||
|
Социальные системы |
Социальные организации |
|
|
Трансцендентные системы или системы, лежащие в настоящий момент вне нашего познания |
Мир живых существ, включая человека, представлен биологическими (живыми) системами различной структурной организации и разного уровня соподчинения, или согласованности. Остановимся на понятии «биологическая система», признаках биологических систем и их уровнях.
Биологические системы – это объекты различной сложности, имеющие несколько уровней структурно-функциональной организации и представляющие собой совокупность взаимосвязанных и взаимодействующих элементов.
Примерами биологических систем являются: клетка, ткани, органы, организмы, популяции, виды, биоценозы, экосистемы разных рангов и биосфера.
Элементарной биологической системой, т.е. системой самого низшего ранга, является клетка, т.к. нет систем еще более низкого ранга, которые бы обладали всей совокупностью признаков, присущих биологическим системам.
Человек занимает особое место среди систем, он не только живет в мире систем, но и сам является системой, персонифицированной составляющей природы.
Гомеостаз
Жизнедеятельность организма, как открытой биосистемы обеспечивается процессами, упорядоченными во времени и пространстве, приуроченными к определённой системе жизнеобеспечения. Эти процессы образуют три потока:
1.информационный 2.поток веществ 3.поток энергии
Вследствие целого ряда сложных химических превращений вещества из окружающей среды уподобляются веществам живого организма и из них строится тело. Эти процессы называют ассимиляцией или пластическим обменом.
С другой стороны сложные органические соединения распадаются на простые, при этом утрачивается их сходство с веществами организма и выделяется энергия, необходимая для реакций биосинтеза. Эти процессы называют диссимиляцией или энергетическим обменом.
Обмен веществ обеспечивает гомеостаз организма.
Эти потоки осуществляются непрерывно и составляют условия существования организма в постоянно меняющейся окружающей среде.
Под руководством нервной и эндокринной систем системы жизнеобеспечения поддерживают процессы внутри организма в оптимальном режиме.
"Гомеостаз – относительное динамическое постоянство состава и свойств внутренней среды, а также устойчивость физиологических функций организма".
К. Бернар Гомеостаз - это способность организма поддерживать основные параметры жизнедеятельности на оптимальном уровне. Важным аспектом гомеостаза организма является стабильность развития (морфогенетический гомеостаз) -способность организма к формированию генетически детерминированного фенотипа при минимальном уровне онтогенетических нарушений.
Также смотрите:
Результаты и обсуждение. Исследование активности карбоксипептидазы N в сыворотке крови онкологических больных при
химиотерапевтическом воздействии
Результаты исследования показали увеличение активности КПN у онкологических больных в период до проведения химиотерапии по сравнению с контрольной группой в 2 раза, и уменьшение активности фермента после проведения химиотерапии, по отношению к периоду до начала лечени...
Бонитировка угодий района по косуле
Хорошие угодья (I бонитета) – угодья, в которых преобладают свойственные и благоприятные для обитания типы угодий. Непригодных для вида нет. Защитно-гнездовые условия хорошие. В таких угодьях имеется обильный и разнообразный набор кормов, устойчивый по годам. Это стац...
Дон - река в Европейской части России. По площади водосбора, равной 422 тыс. км² уступает только Волге, Днепру и Каме. Длина реки - 1870 км. Название происходит от скифо-сарматского dānu, осетинского "дон" вода, река.
Исток Дона расположен в север...
Страница 1 из 2
Рефорат Биологические системы, способы их применения и значение на земле и в космосе
Введение…………………………………………………………………
1. Общая Характеристика работы………………………………
1.1. Цели и задачи……………………………………
1.3. Актуальность темы…………………………………
1.4. Положения, выполняемые на защиту………………
1.5. Структура и объем работы………………………
2. Обзор литературы…………………………
2.1. Биологические системы ………………………………
Что такое биологические системы……………………
История биологических систем………………
2.2. Примеры биологических систем, их существование и значение…
Подземные теплицы – прообраз биосистемы………………………….
Первые шаги к космическим теплицам…………
Микро-биосистемы в квартире…………………………
3. Результаты изучения………………………
3.1. Биологическая система с кактусами – модель космической станции………………………………………
3.2. Круговорот веществ в биологической системе……
Выводы……………………………………………………
Приложения……………………………………………
Список использованных источников………
Введение
С древних времен человечество стремится достичь космоса и его планет. Это возможно только при обеспечении человека всем необходимым (водой, едой, воздухом). Для решения этой задачи необходимы биологические системы, которые будут снабжать экипаж всем необходимым и утилизировать отходы жизнедеятельности.
1. Общая характеристика работы
1.1. Цель и задачи:
· показать многообразие биологических систем, их значение в жизни человека на Земле и космосе;
· показать возможность моделирования биологических систем и их использование;
1.2. Актуальность темы.
На данный момент тема останется актуальной до тех пор, пока ученые не смогут сделать такую биологическую систему, которую можно поместить на космический корабль, чтобы она обеспечивала экипаж всем необходимым и действовала довольно долго.
А также тема актуальна, потому что на Земле во многих странах ощущается недостаток овощей и фруктов. Решить этот вопрос помогут биологические системы, которыми можно занять пустыни, высокогорья и пустые выработки старых шахт. Тем самым удастся решить проблему голода.
1.3. Апробация работы.
Работа доложена на заседании секции биологии МАН Молодежненской ОШ № 1 13.11.2004 года и одобрена к защите.
1.4. Положения, выполняемые на защиту.
· что такое биосистема.
· круговорот веществ биосистеме.
· создание биологической системы с использованием суккулентов.
· значение биологических систем на Земле и в космосе.
1.5. Структура и объем работы.
Работа изложена на ___ страницах, включая ___ рисунка.
2. Обзор литературы.
2.1. Биологические системы
Что такое биологическая система?
Биологическая система – это биологические объекты различной сложности (клетки, ткани, организмы, биоценозы), имеющие, как правило, несколько уровней структурно-функциональной организации. Представляя собой совокупность взаимосвязанных и взаимодействующих элементов, биологические системы обладают свойствами целостности, способности к саморегуляции, что и определяет их устойчивость, а также способность к адаптации по отношению к внешней среде, развитию, самовоспроизведению и эволюции. Любая биологическая система является динамической – в ней постоянно протекает множество процессов, часто сильно различающихся во времени. В то же время биологическая система – открытая система, условием существования которой служит обмен энергией, веществом и информацией как между частями системы, так и с окружающей средой. Важнейшая особенность биологической системы заключается в том, что такой обмен осуществляется под контролем специальных механизмов реализации генетической информации и внутреннего управления, которые позволяют избежать «термодинамической смерти» путем использования энергии, извлекаемой из внешней среды.
Устойчивость стационарных состояний биологической системы (сохранение постоянства внутренних характеристик на фоне нестабильной или изменяющейся внешней среды, а также способность к их переходу из одного состояния в другое, свойство неустойчивости стационарных состояний биологических систем) обеспечиваются многообразными механизмами саморегуляции. В основе саморегуляции биологических систем лежит принцип обратной связи, положительной или отрицательной. Так в цепи регулирования с отрицательной обратной связью, информация об отклонении регулируемой величины от заданного уровня включает в действие регулятор, который действует на регулируемый объект, таким образом, что регулируемая величина возвращается к исходному уровню (знак применения её обратен знаку первоначального отклонения). Этот механизм, а также более сложные комбинации нескольких механизмов могут функционировать на разных уровнях организации биологических систем (например, на молекулярном – ингибирование ключевого фермента при избытке конечного продукта или репрессия синтеза ферментов, на клеточном – гормональная саморегуляция и контрактное угнетение, обеспечивающее оптимальную плотность клеточной популяции, на уровне организмов – регуляция содержания глюкозы в крови, а в общих случаях гомеостаз, обеспечивающих стабильность внутренней среды организма). Специальные механизмы положительной обратной связи (воздействие на регулируемый объект вызывают изменения, совпадающие по знаку с первоначальным отклонением регулируемой величины, вследствие чего система выходит из данного стационарного состояния) лежат в основе перехода биологических систем из одного стационарного состояния в другое и основанных на этих переходах закономерных изменениях биологических систем, обеспечивающих их адаптацию к изменяющимся внешним условиям, перемещение, другие многообразные активные функции биологических систем, их эволюцию. Сложное автономное (независимое от среды) движение биосистем возможно благодаря множественности стационарных состояний биосистем, между которыми могут совершаться переходы. В некоторых случаях новое состояние оказывается не стационарным, а автоколебательным, т. е. таким, в котором значения показателей колеблются во времени с постоянной амплитудой. Такие явления в основе периодических процессов в биологических системах временной организации биосистем, в основе функционирования биологических часов.
При анализе поведения и свойств биосистем широкое применение находят различные методы физического и математического моделирования, используются кибернетические и термодинамические подходы. Системный подход оказывается перспективным для решения многих практически важных проблем (таких, например, как создание замкнутых биосистем жизнеобеспечения, проблема заболеваний, связанных с нарушением гомеостаза и прочее). Примером биологической системы являются все живые организмы, населяющие нашу планету, в том числе и растения.
История биологических систем.
Планета Земля в космосе – пример частично закрытой, автономной биологической системы. Живые объекты на Земле обеспечены всеми необходимыми веществами для жизни и жизнедеятельности. Эта система довольно постоянна, получая из космоса главный фактор жизни – солнечный свет. Эта система – Земля, удерживает на своей поверхности и в ближайшем окружении не только тяжелые твердые предметы, но и воздух, воду, пары воды. Самую простейшую модель биологической системы придумали японцы. В Японии такие системы появились на прилавках в восьмидесятые годы двадцатого века. Они были предназначены для занятых или ленивых цветоводов, которые хотели иметь у себя дома отличные сорта цветов, но, в то же время, не ухаживать за ними. Достаточно было подойти к прилавку, купить такую систему, налить туда воды, герметически закрыть и готово – отличные растения у себя дома целый год.
2.2. Примеры биологических систем, их существование и значение.
Подземные теплицы – прообраз биосистемы.
В настоящее время расширение площадей защищенного грунта сдерживается из-за высокой стоимости строительства теплиц и их эксплуатации. Так, в степной зоне Украины строительство 1 м2 теплиц в среднем обходилось в 60 рублей, а в Якутске, где овощи и фрукты в дефиците, 250 рублей. И, как это ни парадоксально, тепличное хозяйство более всего развито в центральных и южных районах Украины, а вовсе не там, где в свежих овощах испытывается недостаток. Проведенные за рубежом опыты показали, что овощи можно с успехом выращивать под землей, используя для этого подземные выработки старых шахт, оставшиеся после добычи полезных ископаемых.
Но не все шахтные выработки пригодны для создания подземных теплиц, т. к. большинство угольных шахт подтапливаются грунтовыми водами и в них наблюдаются выбросы метана и других горючих газов, что значительно удорожает эксплуатацию таких выработок (искусственная откачка грунтовых вод). Положительным моментом подземных выработок является постоянная положительная температура выше +11 оС. Электрические осветительные приборы и оборудование выделяют дополнительное тепло и ещё подогревают шахтные выработки и температура в них даже зимой при сильных морозах достаточна для выращивания большинства овощных растений. В более глубоких шахтах с глубиной температура повышается, но увеличиваются затраты на вертикальную транспортировку людей, оборудования и продукции, а также откачку грунтовых вод и вентиляцию.
Под землей растения можно выращивать круглый год. Традиционные культивационные сооружения (теплицы, парники, оранжереи) зимой приходится обогревать, а летом охлаждать, но следует еще и вентилировать их, на что тратится 20-30% всей потребляемой энергии. Наличие в Украине дешевой электрической энергии для освещения, постоянная температура окружающих пород на глубине и низкая стоимость подготовки отработанных шахт к производству сельскохозяйственной продукции могут дать большой эффект. Что необходимо для нормального роста растений в искусственных условиях? Прежде всего освещение и среда обитания корней – субстрат.
Опыт эксплуатации подземных выработок в Криворожском железорудном бассейне показал: применяя искусственное освещение соответствующего спектрального состава, можно получать очень хороший урожай овощей. В качестве источников света здесь применяются облучатели тепличные ОТ-400, лампы РРЛ-700 и лампы накаливания мощностью 500 и 1000 Вт. Если позволят габариты помещения, можно использовать и другие системы искусственного освещения, установку ускоренного выращивания растений (УВР), установку системы освещения растительных теплиц СОРТ 1-5 и СОРТ 1-10, систему освещения вегетационных теплиц СОВК-1 и другие (такие установки использовались и на подводных лодках).
В шахтах наиболее традиционно применять гидропонный способ выращивания растений. Кроме того, субстраты (гравий, торф, шлак, песок), соответственно подобранные по физиологическим и химическим свойствам, могут «работать» в подземных условиях 10-15 лет и более, тогда как почву в теплицах необходимо менять каждые 3-4 года.
При выращивании овощей в подземных теплицах на гидропонике многие отрицательные свойства почвы (необходимость внесения большого количества удобрений, повышенная влажность, наличие вредителей и возбудителей болезней растений) полностью отсутствуют. Соотношение питательных веществ в растворах может быть изменено в зависимости от возраста растений и времени созревания плодов, что значительно повышает коэффициент полезного действия вносимых удобрений и урожай возделываемых культур. Созревание овощей при этих условиях происходит среднем на 2-3 недели раньше, чем в грунтовых теплицах.
При подземном выращивании овощей лотки для гидропоники могут быть изготовлены из различных материалов – дерева, обернутого пленкой, пластмассы или асбоцементных труб большого диаметра. Лотки должны укладываться с уклоном 0,3о для стока питательного раствора.
Гидропоника способствует лучшему обеспечению овощей водой с растворенными в ней питательными веществами, предохраняет зону расположения корней от колебания температуры и подсыхания, позволяет автоматизировать производственные процессы по уходу за растениями, а также значительно повысить урожайность продукции.
В настоящее время поступает все больше и больше сообщений об использовании шахтных выработок для выращивания овощей – в Казахстане, Норильске и в других городах мира взялись за это дело с интересом.
Актуальность же проблемы не вызывает сомнений – подземные теплицы должны стать составной частью агропромышленных комплексов, продовольственной программы Украины.
Первые шаги на пути к космическим теплицам.
Сложнее по устройству, чем подземные теплицы, но более совершенные наземные тепличные комплексы приближаются по конструкции к закрытой биосистеме, получающей извне только солнечную энергию и информационные сигналы связи. В таких комплексах добровольцы в команде, имитирующей команду космического корабля, проживали и работали более чем по году, изучались вопросы круговорота веществ, в том числе и воды, тепловой баланс, вопросы питания группы людей, реутилизации отходов и другие наблюдения и исследования растений и людей в такой системе. Такие научные комплексы создавались в горах, т. к. условия освещенности (инсоляции) и суточного колебания температур ближе к условиям космического пространства, а на поверхности земли на небольших высотах над уровнем моря условия очень отличаются. На больших высотах, как и в космосе, значительно усилена ультрафиолетовая составляющая солнечной радиации. Также на больших высотах значительно меньше атмосферное давление и наблюдаются резкие колебания температуры за счет сильно меняющегося в течение суток потока солнечной энергии.
Для нормального обеспечения людей, находящихся в таком комплексе, применялись специальные стекла в покрытиях тепличного комплекса. Такое стекло поглощает и ослабляет поток ультрафиолетовой радиации и частично снижает резкие колебания температуры внутри комплекса, т. к. препятствует прохождению не только ультрафиолетовых, но и инфракрасных (тепловых) лучей. Дней поступает меньше тепла, теплица не перегревается, а ночью меньше теряется тепла, теплица не переохлаждается. Для снижения колебания температур применяются также шторки-жалюзи, а также придумана оригинальная система суточных колебаний температуры – это тепловые аккумуляторы. Они используют высокие показатели теплоемкости воды. Черные трубы, наполненные водой, прокладываются на поверхности почвы или субстрата для растений. Днем, при избытке тепла, эти аккумуляторы поглощают тепло, препятствуя перегреву, а ночью отдают его, предохраняя систему от переохлаждения.
Группы людей прожили год и более в помещении теплицы, биологически абсолютно изолированном от атмосферы Земли. Воздух и вода очищались и восстанавливались растениями. Так были сделаны первые шаги человека к созданию надежной замкнутой биологической системы обеспечения человека с использованием высших растений. Эксперимент оказался удачным.
Главная задача при построении космических теплиц состоит в том, чтобы отобрать растения, которые обеспечили бы космонавтов наиболее полезным набором питательных веществ. Поэтому исследователи считают, что с большим успехом эту задачу удастся решить, если на космическом «огороде» посадить набор различных овощей. Только так можно обеспечить полноценную пищу. Естественно, что, подбирая овощи для космической теплицы, необходимо учитывать привычки и вкусы людей. К примеру, зарубежные ученые предлагают выращивать в космической оранжерее бобы, горох и масличное дерево – земляной орех, которые в сочетании с другими растениями удовлетворительно обеспечивают рацион космонавтов. Все это так, но земляной орех для нас непривычная еда. Поэтому рацион космонавтов целесообразно составлять из растений, которые возделываются на территории страны или введены в меню овощей и фруктов, ставших для нас уже привычными. Рацион космонавта не должен включать одни овощи, в нем обязательно должны быть животные белки и жиры. Для решения этого вопроса было рассмотрено большое количество самых различных видов организмов, в том числе беспозвоночных и рыб. Установлено, что современный аквариум – это тоже прообраз замкнутой биоэкологической системы, который можно использовать на корабле. Определено, что на борту космического корабля должны быть и травоядные животные. Более всего для обеспечения космонавтов мясом подходят куры. Они едят практически все и легко размножаются.
Раздел 1.2. Уровневая организация и
эволюция. Основные уровни организации живой природы: клеточный,
организменный, популяционно-видовой, биогеоценотический, биосферный.
Биологические системы. Общие признаки биологических систем: клеточное
строение, особенности химического состава, обмен веществ и превращения
энергии, гомеостаз, раздражимость, движение, рост и развитие,
воспроизведение, эволюция.
Основные термины и понятия, проверяемые в экзаменационных работах: уровень
жизни, биологические системы, изучаемые на данном уровне,
молекулярно-генетический, клеточный, организменный, популяционно –
видовой, биогеоценотический, биосферный.
Уровни организации живых систем отражают соподчиненность, иерархичность структурной организации жизни. Уровни жизни отличаются друг от друга сложностью организации системы. Клетка устроена проще по сравнению с многоклеточным организмом или популяцией.
Уровень жизни – это форма и способ ее существования. Например, вирус существует в виде молекулы ДНК или РНК, заключенной в белковую оболочку. Это форма существования вируса. Однако свойства живой системы вирус проявляет, только попав в клетку другого организма. Там он размножается. Это способ его существования.
Молекулярно-генетический уровень представлен отдельными биополимерами (ДНК, РНК, белками, липидами, углеводами и другими соединениями); на этом уровне жизни изучаются явления, связанные с изменениями (мутациями) и воспроизведением генетического материала, обменом веществ.
Клеточный – уровень, на котором жизнь существует в форме клетки – структурной и функциональной единицы жизни. На этом уровне изучаются такие процессы, как обмен веществ и энергии, обмен информацией, размножение, фотосинтез, передача нервного импульса и многие другие.
Организменный – это самостоятельное существование отдельной особи – одноклеточного или многоклеточного организма.
Популяционно-видовой – уровень, который представлен группой особей одного вида – популяцией; именно в популяции происходят элементарные эволюционные процессы – накопление, проявление и отбор мутаций.
Биогеоценотический – представлен экосистемами, состоящими из разных популяций и среды их обитания.
Биосферный – уровень, представляющий совокупность всех биогеоценозов. В биосфере происходит круговорот веществ и превращение энергии с участием организмов. Продукты жизнедеятельности организмов участвуют в процессе эволюции Земли.
Основные термины и понятия, проверяемые в экзаменационной работе: гомеостаз, единство живой и неживой природы, изменчивость, наследственность, обмен веществ.
Признаки и свойства живого . Живые системы имеют общие признаки:
Клеточное строение – все существующие на Земле организмы состоят из клеток. Исключением являются вирусы, проявляющие свойства живого только в других организмах.
Обмен веществ – совокупность биохимических превращений, происходящих в организме и других биосистемах.
Саморегуляция – поддержание постоянства внутренней среды организма (гомеостаза). Стойкое нарушение гомеостаза ведет к гибели организма.
Раздражимость – способность организма реагировать на внешние и внутренние раздражители (рефлексы у животных и тропизмы, таксисы и настии у растений).
Изменчивость – способность организмов приобретать новые признаки и свойства в результате влияния внешней среды и изменений наследственного аппарата – молекул ДНК.
Наследственность – способность организма передавать свои признаки из поколения в поколение.
Репродукция или самовоспроизведение – способность живых систем воспроизводить себе подобных. В основе размножения лежит процесс удвоения молекул ДНК с последующим делением клеток.
Рост и развитие – все организмы растут в течение своей жизни; под развитием понимают как индивидуальное развитие организма, так и историческое развитие живой природы.
Открытость системы – свойство всех живых систем связанное с постоянным поступлением энергии извне и удалении продуктов жизнедеятельности. Иными словами организм жив, пока в нем происходит обмен веществами и энергией с окружающей средой.
Способность к адаптациям – в процессе исторического развития и под действием естественного отбора организмы приобретают приспособления к условиям окружающей среды (адаптации). Организмы, не обладающие необходимыми приспособлениями, вымирают.
Общность химического состава . Главными особенностями химического состава клетки и многоклеточного организма являются соединения углерода – белки, жиры, углеводы, нуклеиновые кислоты. В неживой природе эти соединения не образуются.
Общность химического состава живых систем и неживой природы говорит о единстве и связи живой и неживой материи. Весь мир представляет собой систему, в основании которой лежат отдельные атомы. Атомы, взаимодействуя друг с другом, образуют молекулы. Из молекул в неживых системах формируются кристаллы горных пород, звезды, планеты, вселенная. Из молекул, входящих в состав организмов формируются живые системы – клетки, ткани, организмы. Взаимосвязь живых и неживых систем отчетливо проявляется на уровне биогеоценозов и биосферы.