Reticulul endoplasmatic este neted. Reticul endoplasmatic neted
14. Eps granulat și neted. Structura și caracteristicile funcționării în celule de același tip.
Reticulul endoplasmatic (EPS) - un sistem de canale tubulare comunicante sau separate și cisterne aplatizate situate în întreaga citoplasmă a celulei. Sunt delimitate de membrane (organite membranare). Uneori, rezervoarele au expansiuni sub formă de bule. Canalele EPS se pot conecta cu membrane de suprafață sau nucleare, contact cu complexul Golgi.
Reticulul endoplasmatic este o structură spațială fragmentată delimitată de o membrană simplă. În imediata apropiere a miezului, formează un sistem de cisterne plate și subțiri, cu distanța crescândă față de miez, cisternele devin mai groase până când în cele din urmă devin tubulare. Spre membrana celulară, diametrul tuburilor tinde să scadă, iar tuburilor le lipsește complet ribozomul asociat cu suprafața lor. Aspectul, locația și numărul de rezervoare sau tuburi depind de activitatea metabolică a celulei.
În acest sistem se pot distinge EPS neted și aspru (granular).
EPS dur. Pe canalele ER rugoase, ribozomii sunt localizați sub formă de polizomi. Aici are loc sinteza proteinelor, produse în principal de celulă pentru export (scoaterea din celulă), de exemplu, secreții de celule glandulare. Aici are loc formarea lipidelor și proteinelor membranei citoplasmatice și asamblarea acestora. Cisternele strâns împachetate și canalele ER granulare formează o structură stratificată în care sinteza proteinelor este cea mai activă. Acest loc se numește ergastoplasmă.
În zona de tranziție, există trotuare sub formă de rezervoare și tubuli groși, care sunt conectate prin ribozomi la suprafața lor și sunt complet lipsite de ribozomi. Tuburile sale sunt literalmente împletite cu un alt corp celular, iar acesta este aparatul Golgi. Membrana reticulului endoplasmatic este foarte asemănătoare structural cu membrana celulară, dar este mai subțire. Analizând prezența acidului ribonucleic în această membrană, proprietățile sale diferă de alți acizi ribonucleici conținuti în celulă.
Reticulul endoplasmatic aspru
De fapt, membrana nucleară exterioară poate fi considerată ca fiind membrana reticulului endoplasmatic sever. Suprafața acestei membrane este dens acoperită cu ribozomi. Cel mai adesea în desenele schematice, acesta este ilustrat ca un sistem de rezervoare plate și înguste care încapsulează nucleul celulei. De fapt, însă, rezervoarele pot fi chiar amplasate perpendicular pe miez.
EPS neted. Nu există ribozomi pe membranele netede ale RE. Aici are loc în principal sinteza grăsimilor și substanțelor similare (de exemplu, hormoni steroizi), precum și a carbohidraților. Prin canalele EPS netede, materialul finit se deplasează, de asemenea, la locul de ambalare în granule (în zona complexului Golgi). În celulele hepatice, ER neted participă la distrugerea și neutralizarea unui număr de substanțe toxice și medicinale (de exemplu, barbiturice). În mușchii striați, tubii și cisternele ER netede depun ioni de calciu.
15. Complexul Golgi. Structură și funcții.
Complexul Golgi este o structură membranară inerentă oricărei celule eucariote. Complexul Golgi este format din cisterne aplatizate, de obicei dispuse în stive (dictiozomi). Rezervoarele nu sunt izolate, ci sunt interconectate printr-un sistem de tuburi. Prima cisternă din nucleu se numește cis-pol al complexului Golgi, iar ultima, respectiv, trans-pol. Numărul de cisterne din diferite celule ale diferitelor organisme poate varia, dar, în general, structura complexului Golgi în toate eucariotele este aproximativ aceeași. În celulele secretoare, este deosebit de puternic dezvoltat. Funcțiile complexului Golgi sunt de a transporta proteinele la destinație, precum și de glicozilare, deglicozilare și modificarea lanțurilor de oligozaharide.
Complexul Golgi este caracterizat de anizotropie funcțională. Proteinele nou sintetizate sunt transportate din reticulul endoplasmatic la polul cis al dictiozomilor folosind vezicule. Apoi se deplasează treptat spre trans-pol, suferind modificări treptate (pe măsură ce se îndepărtează de nucleu, compoziția sistemelor enzimatice din rezervoare se modifică). În cele din urmă, proteinele călătoresc la destinația lor finală în vezicule care înmugurează din polul trans. Complexul Golgi asigură transportul proteinelor în trei compartimente: la lizozomi (precum și la vacuola centrală). celula planteiși vacuolele contractile ale protozoarelor), către membrana celulară și în spațiul intercelular. Direcția transferului de proteine este determinată de etichete glicozidice speciale. De exemplu, un marker pentru enzimele lizozomale este manoza-6-fosfat. Maturarea și transportul proteinelor mitocondriale, nucleare și cloroplastice au loc fără participarea complexului Golgi: ele sunt sintetizate de ribozomi liberi și apoi intră direct în citosol. O funcție importantă a complexului Golgi este sinteza și modificarea componentei carbohidrate a glicoproteinelor, proteoglicanilor și glicolipidelor. De asemenea, sintetizează multe polizaharide, cum ar fi hemiceluloza și pectina din plante. Cisternele complexului Golgi conțin o gamă întreagă de glicoziltransferaze și glicozidaze diferite. Ele suferă, de asemenea, sulfatarea reziduurilor de carbohidrați.
Celulele, care este un sistem ramificat de cavități aplatizate, înconjurate de o membrană, vezicule și tubuli.
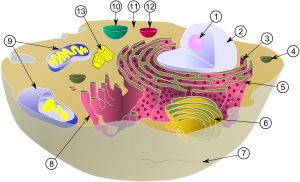
Reprezentare schematică nucleul celular, reticulul endoplasmatic și complexul Golgi.
(1) Nucleul celular.
(2) Porii membranei nucleare.
(3) Reticulul endoplasmatic granular.
(4) Reticulul endoplasmatic agranular.
(5) Ribozomi de pe suprafața reticulului endoplasmatic granular.
(6) Proteine transportate.
(7) Vezicule de transport.
(8) Complexul Golgi.
(9)
(10)
(11)
Istoria descoperirilor
Pentru prima dată, reticulul endoplasmatic a fost descoperit de omul de știință american C. Porter în 1945 folosind microscopia electronică.
Structura
Reticulul endoplasmatic este format dintr-o rețea extinsă de tubuli și pungi înconjurate de o membrană. Aria membranelor reticulului endoplasmatic este mai mult de jumătate din suprafața totală a tuturor membranelor celulare.
Membrana ER este morfologic identică cu învelișul nucleului celular și este una cu aceasta. Astfel, cavitățile reticulului endoplasmatic se deschid în cavitatea intermembranară a membranei nucleare. Membranele EPS asigură transportul activ al unui număr de elemente împotriva unui gradient de concentrație. Filamentele care formează reticulul endoplasmatic au un diametru de 0,05-0,1 µm (uneori până la 0,3 µm), grosimea membranelor cu două straturi care formează peretele tubilor este de aproximativ 50 angstromi (5 nm, 0,005 µm). Aceste structuri conțin fosfolipide nesaturate, precum și unele colesterol și sfingolipide. Conțin și proteine.
Tubulii, al căror diametru variază de la 0,1-0,3 µm, sunt umpluți cu conținut omogen. Funcția lor este implementarea comunicării între conținutul veziculelor EPS, mediul extern și nucleul celular.
Reticulul endoplasmatic nu este o structură stabilă și este supus unor modificări frecvente.
Există două tipuri de EPR:
- reticul endoplasmatic granular
- reticul endoplasmatic agranular (neted).
Pe suprafața reticulului endoplasmatic granular se află un numar mare de ribozomi care sunt absenți de pe suprafața RE agranulară.
Reticulul endoplasmatic granular și agranular îndeplinesc diferite funcții în celulă.
Funcțiile reticulului endoplasmatic
Cu participarea reticulului endoplasmatic, au loc translația și transportul proteinelor, sinteza și transportul lipidelor și steroizilor. EPS se caracterizează și prin acumularea de produse de sinteză. Reticulul endoplasmatic este, de asemenea, implicat în crearea unei noi membrane nucleare (de exemplu, după mitoză). Reticulul endoplasmatic conține o cantitate intracelulară de calciu, care este, în special, un mediator al contracției celulelor musculare. În celulele fibrelor musculare există o formă specială a reticulului endoplasmatic - reticulul sarcoplasmic.
Funcțiile reticulului endoplasmatic agranular
Reticulul endoplasmatic agranular este implicat în multe procese metabolice. Enzimele reticulului endoplasmatic agranular sunt implicate în sinteza diferitelor lipide și fosfolipide, acizi grași și steroizi. De asemenea, reticulul endoplasmatic agranular joaca un rol important in metabolismul carbohidratilor, dezinfectarea celulelor si depozitarea calciului. În special, în legătură cu aceasta, reticulul endoplasmatic agranular predomină în celulele glandelor suprarenale și ficatului.
Sinteza hormonilor
Hormonii care se formează în EPS agranular includ, de exemplu, hormoni sexuali de vertebrate și hormoni steroizi suprarenali. Celulele testiculare și ovariene responsabile de sinteza hormonală conțin cantități mari de reticul endoplasmatic agranular.
Acumularea și conversia carbohidraților
Carbohidrații din organism sunt stocați în ficat sub formă de glicogen. Glicoliza transformă glicogenul din ficat în glucoză, care este un proces critic în menținerea nivelului de glucoză din sânge. Una dintre enzimele EPS agranulare desprinde o grupare fosfo din primul produs al glicolizei, glucoza-6-fosfat, permițând astfel glucozei să părăsească celula și să crească nivelul zahărului din sânge.
Neutralizarea otrăvurilor
Reticulul endoplasmatic neted al celulelor hepatice Participarea activăîn neutralizarea tot felul de otrăvuri. Enzimele netede ER ataseaza moleculele de substante active intalnite, care pot fi astfel dizolvate mai rapid. În cazul aportului continuu de otrăvuri, droguri sau alcool se formează o cantitate mai mare de EPR agranular, care crește doza de substanță activă necesară pentru a obține același efect.
Reticulul sarcoplasmic
O formă specială a reticulului endoplasmatic agranular, reticulul sarcoplasmatic, formează ER în celulele musculare, în care ionii de calciu sunt pompați activ din citoplasmă în cavitatea ER împotriva gradientului de concentrație în starea neexcitată a celulei și eliberați în citoplasmă. pentru a iniția contracția. Concentrația ionilor de calciu în EPS poate ajunge la 10 −3 moli, în timp ce în citosol este de aproximativ 10 −7 moli (în repaus). Astfel, membrana reticulului sarcoplasmatic asigură transport activ împotriva gradienților de concentrație de ordin înalt. Iar aportul și eliberarea ionilor de calciu în EPS este într-o relație subtilă cu condițiile fiziologice.
Concentrația ionilor de calciu în citosol afectează multe procese intracelulare și intercelulare, cum ar fi: activarea sau inhibarea enzimelor, expresia genelor, plasticitatea sinaptică a neuronilor, contracțiile celulelor musculare, eliberarea de anticorpi din celulele sistemului imunitar.
Funcțiile reticulului endoplasmatic granular
Reticulul endoplasmatic granular are două funcții: sinteza proteinelor și producerea membranei.
Sinteza proteinei
Proteinele produse de celulă sunt sintetizate pe suprafața ribozomilor, care pot fi atașați la suprafața RE. Lanțurile polipeptidice rezultate sunt plasate în cavitatea reticulului endoplasmatic granular (unde cad și lanțurile polipeptidice sintetizate în citosol), unde sunt ulterior tăiate și pliate în mod corect. Astfel, secvențele liniare de aminoacizi se obțin după translocarea în reticulul endoplasmatic a structurii tridimensionale necesare, după care sunt retransferate în citosol.
Sinteza membranei
Ribozomii atașați la suprafața ER granulară produc proteine, care, împreună cu producția de fosfolipide, printre altele, extinde suprafața membranei proprii a ER, care trimite fragmente de membrană către alte părți ale sistemului membranar prin vezicule de transport.
Vezi si
- Reticulonii sunt proteine ale reticulului endoplasmatic.
Fundația Wikimedia. 2010 .













