Reticulul endoplasmatic. Organele de importanță generală. Reticulul endoplasmatic
14. Eps granulat și neted. Structura și caracteristicile funcționării în celule de același tip.
Reticulul endoplasmatic (EPS) - un sistem de canale tubulare comunicante sau separate și cisterne aplatizate situate în întreaga citoplasmă a celulei. Sunt delimitate de membrane (organite membranare). Uneori, rezervoarele au expansiuni sub formă de bule. Canalele EPS se pot conecta cu membrane de suprafață sau nucleare, contact cu complexul Golgi.
În acest sistem se pot distinge EPS neted și aspru (granular).
EPS dur. Pe canalele ER rugoase, ribozomii sunt localizați sub formă de polizomi. Aici are loc sinteza proteinelor, produse în principal de celulă pentru export (scoaterea din celulă), de exemplu, secreții de celule glandulare. Aici are loc formarea lipidelor și proteinelor membranei citoplasmatice și asamblarea acestora. Cisternele strâns împachetate și canalele ER granulare formează o structură stratificată în care sinteza proteinelor este cea mai activă. Acest loc se numește ergastoplasmă.
EPS neted. Nu există ribozomi pe membranele netede ale RE. Aici are loc în principal sinteza grăsimilor și substanțelor similare (de exemplu, hormoni steroizi), precum și a carbohidraților. Prin canalele EPS netede, materialul finit se deplasează, de asemenea, la locul de ambalare în granule (în zona complexului Golgi). În celulele hepatice, ER neted participă la distrugerea și neutralizarea unui număr de substanțe toxice și medicinale (de exemplu, barbiturice). În mușchii striați, tubii și cisternele ER netede depun ioni de calciu.
15. Complexul Golgi. Structură și funcții.
Complexul Golgi este o structură membranară inerentă oricărei celule eucariote. Complexul Golgi este format din cisterne aplatizate, de obicei dispuse în stive (dictiozomi). Rezervoarele nu sunt izolate, ci sunt interconectate printr-un sistem de tuburi. Prima cisternă din nucleu se numește cis-pol al complexului Golgi, iar ultima, respectiv, trans-pol. Numărul de cisterne din diferite celule ale diferitelor organisme poate varia, dar, în general, structura complexului Golgi în toate eucariotele este aproximativ aceeași. În celulele secretoare, este deosebit de puternic dezvoltat. Funcțiile complexului Golgi sunt de a transporta proteinele la destinație, precum și de glicozilare, deglicozilare și modificarea lanțurilor de oligozaharide.
Complexul Golgi este caracterizat de anizotropie funcțională. Proteinele nou sintetizate sunt transportate din reticulul endoplasmatic la polul cis al dictiozomilor folosind vezicule. Apoi se deplasează treptat spre trans-pol, suferind modificări treptate (pe măsură ce se îndepărtează de nucleu, compoziția sistemelor enzimatice din rezervoare se modifică). În cele din urmă, proteinele călătoresc la destinația lor finală în vezicule care înmugurează din polul trans. Complexul Golgi asigură transportul proteinelor în trei compartimente: către lizozomi (precum și către vacuola centrală a celulei vegetale și vacuolele contractile ale protozoarelor), către membrana celulară și către spațiul intercelular. Direcția transferului de proteine este determinată de etichete glicozidice speciale. De exemplu, un marker pentru enzimele lizozomale este manoza-6-fosfat. Maturarea și transportul proteinelor mitocondriale, nucleare și cloroplastice au loc fără participarea complexului Golgi: ele sunt sintetizate de ribozomi liberi și apoi intră direct în citosol. O funcție importantă a complexului Golgi este sinteza și modificarea componentei carbohidrate a glicoproteinelor, proteoglicanilor și glicolipidelor. De asemenea, sintetizează multe polizaharide, cum ar fi hemiceluloza și pectina din plante. Cisternele complexului Golgi conțin o gamă întreagă de glicoziltransferaze și glicozidaze diferite. Ele suferă, de asemenea, sulfatarea reziduurilor de carbohidrați.
Reticul endoplasmatic (reticul endoplasmatic)- un organel al unei celule eucariote. Sub forma unei rețele de canale și rezervoare, limitate de o singură membrană, se ramifică pe întregul volum al citoplasmei. Reticulul endoplasmatic este implicat în metabolism: sintetizează lipide pentru membrana dublă exterioară a celulei și pentru membrana proprie, unică, asigură transportul de substanțe între organele celulare, servește drept tezaur de substanțe și loc pentru izolarea acestora.
Există două tipuri de reticul endoplasmatic - aspru și neted. Rețeaua aspră poartă numeroși ribozomi pe suprafața exterioară. Proteinele sintetizate pe ele sunt izolate aici de alte proteine ale celulei prin transferul lor prin membrana canalului reticulului endoplasmatic. Ea „recunoaște proteinele trecute după” semnalul lor special „termină. Scindarea acestor capete după trecerea proteinei prin membrană se numește maturare proteică. Unele proteine – secretoare – sunt eliberate din celulă. Altele sunt incluse în toate celulele. membranelor.
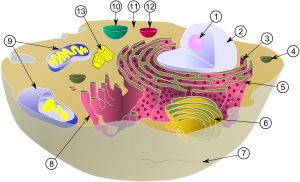
Reprezentare schematică nucleul celular, reticulul endoplasmatic și complexul Golgi.
(1) Nucleul celular. (2) Porii membranei nucleare. (3) Reticulul endoplasmatic granular.
(4) Reticulul endoplasmatic agranular. (5) Ribozomi de pe suprafața granularului
reticulul endoplasmatic. (6) Macromolecule (7) Vezicule de transport.
(8) Complexul Golgi. (9) Cis-Golgi (10) Trans-Golgi (11) Cisterne Golgi
O rețea netedă constă din tuburi, canale și bule cu o secțiune transversală mai mică decât într-o rețea brută. Funcțiile sale sunt, de asemenea, diverse: aici se sintetizează lipidele membranare, dar, pe lângă acestea, lipidele nemembranare (de exemplu, hormoni speciali animale), substanțele toxice sunt neutralizate de complexe enzimatice speciale, iar ionii se acumulează. Deci, în mușchii striați, rețeaua netedă servește ca un rezervor de ioni de calciu. Membranele acestei rețele conțin „pompe” puternice de calciu care, în sutimi de secundă, sunt transferate în orice direcție un numar mare de ionii de calciu. În celulele specializate, aspectul unei rețele netede este diferit, care este asociat cu funcțiile sale specifice în metabolismul intracelular.
Reticulul endoplasmatic este foarte vulnerabil la impact: pierde rapid ribozomii și se prăbușește. Cu toate acestea, datorită capacității de a reconstrui rapid, se poate recupera.
1. Cărui grup de organite aparțin lizozomii, reticulul endoplasmatic si aparatul Golgi?
Ele aparțin organelelor cu o singură membrană.
2. Care este structura și funcția reticulului endoplasmatic? Cum este EPS aspru diferit de neted?
Reticulul endoplasmatic (EPS) sau reticulul endoplasmatic (ER), este un sistem de canale și cavități înconjurate de o membrană și care pătrund în hialoplasma celulei. Distingeți între EPS aspru și neted. Reticulul endoplasmatic dur conține mulți ribozomi. Aici sunt sintetizate proteinele care sunt excretate în afara celulei. Proteinele membranare sunt de asemenea sintetizate pe ribozomii ER brut. Sinteza carbohidraților și a lipidelor are loc pe suprafața EPS neted. În plus, ionii de calciu, care sunt regulatori importanți ai funcțiilor celulare și ale corpului în ansamblu, se acumulează în ER neted. Substanțele care se formează pe membranele EPS se acumulează în interiorul cavităților rețelei și sunt transformate. De exemplu, proteinele capătă structura lor caracteristică secundară, terțiară sau cuaternară. EPS dur este mai bine dezvoltat în acele celule care sintetizează o cantitate mare de proteine (de exemplu, în celulele glandelor salivare și pancreasului, care realizează sinteza enzimelor digestive; în celulele pancreasului și glandei pituitare). Smooth ER este bine dezvoltat în celulele care sintetizează, de exemplu, polizaharide și lipide (celule ale glandelor suprarenale și ale gonadelor care produc hormoni steroizi; celule hepatice care sintetizează glicogen etc.)
3. Cum funcționează complexul Golgi? Ce funcții îndeplinește?
Complexul (aparatul) Golgi este un sistem de structuri membranare intracelulare: rezervoare și vezicule, în care se acumulează substanțe sintetizate pe membranele EPS. Substanțele sunt livrate în complexul Golgi în vezicule membranoase care se desprind de reticulul endoplasmatic și se unesc cu cisternele complexului Golgi. Aici, aceste substanțe suferă diferite transformări biochimice și apoi sunt din nou împachetate în vezicule membranare, iar cele mai multe dintre ele sunt transportate în membrana citoplasmatică. Membrana veziculelor fuzionează cu membrana citoplasmatică, iar conținutul este expulzat în afara celulei prin exocitoză. În complexul Golgi celule vegetale se sintetizează polizaharidele din peretele celular (cochilii). O altă funcție importantă a complexului Golgi este formarea lizozomilor.
4. Majoritatea complexe mari Golgi (până la 10 microni) se găsesc în celulele glandelor endocrine. Ce crezi, cu ce este legat?
Glandele endocrine produc hormoni care sunt necesari proceselor vitale ale organismului. Aceeași formare a unui număr de hormoni și maturarea lor se realizează în cavitățile complexului Golgi.
5. Ce este comun în structura și funcțiile reticulului endoplasmatic și complexului Golgi? Care este diferența?
Similitudine: sunt organite cu o singură membrană și reprezintă un sistem de cavități și cisterne. Diferențe: în EPS are loc formarea de substanțe, care apoi intră în complexul Golgi și suferă transformări biochimice (se coace). De asemenea, complexul Golgi este implicat în formarea lizozomilor.
6. Ce sunt lizozomii? Cum sunt formate? Ce funcții îndeplinesc?
Lizozomii sunt mici vezicule membranoase care se împletesc din cisternele complexului Golgi și conțin un set de enzime digestive. Enzimele lizozomilor sunt capabile să descompună proteinele, carbohidrații, lipidele, acizi nucleici. Pe lângă digerarea substanțelor care intră în celulă din exterior, lizozomii participă la descompunerea componentelor interne ale celulei (molecule și organite întregi), deteriorate sau învechite. Acest proces se numește autofagie.
7. Sugerați de ce enzimele din lizozom nu își scindează propria membrană. Care sunt consecințele pentru celula care poate duce la ruperea membranelor lizozomului?
Lizozomii conțin un set extins de enzime hidrolitice capabile să descompună aproape toate tipurile de macromolecule biologice. În mod normal, aceste enzime nu acționează asupra componentelor propriilor celule, deoarece sunt separate de o membrană lizozomală. Când membrana lizozomului se rupe, enzimele din aceasta intră în celulă și încep să absoarbă și să digere conținutul său intern, ceea ce duce la moartea celulei.
8. S-a stabilit că anumite oligo- sau polizaharide sunt „atașate” de moleculele multor substanțe pentru a fi îndepărtate din celulă din complexul Golgi, iar diferite componente glucide sunt atașate la diferite substanțe. În această formă modificată, substanțele sunt excretate în mediul extracelular. De ce crezi că este nevoie de asta?
maturizarea are loc în complexul Golgi diverse substante, care sunt apoi trimise la diferite organite ale celulei sau îndepărtate din aceasta. Un astfel de proces treptat trebuie cumva controlat. Prin urmare, substanțele maturizate sunt marcate cu oligo- și polizaharide speciale, care joacă rolul unui fel de marcă de calitate. Cu ajutorul acestora, substanțele care au trecut prin aparatul Golgi sunt livrate la adresa, în funcție de etichetele pe care le-au primit în aparatul Golgi.
Celulele, care este un sistem ramificat de cavități aplatizate, înconjurate de o membrană, vezicule și tubuli.
Reprezentare schematică a nucleului celular, reticulului endoplasmatic și complexului Golgi.
(1) Nucleul celular.
(2) Porii membranei nucleare.
(3) Reticulul endoplasmatic granular.
(4) Reticulul endoplasmatic agranular.
(5) Ribozomi de pe suprafața reticulului endoplasmatic granular.
(6) Proteine transportate.
(7) Vezicule de transport.
(8) Complexul Golgi.
(9)
(10)
(11)
Istoria descoperirilor
Pentru prima dată, reticulul endoplasmatic a fost descoperit de omul de știință american C. Porter în 1945 folosind microscopia electronică.
Structura
Reticulul endoplasmatic este format dintr-o rețea extinsă de tubuli și pungi înconjurate de o membrană. Aria membranelor reticulului endoplasmatic este mai mult de jumătate din suprafața totală a tuturor membranelor celulare.
Membrana ER este morfologic identică cu învelișul nucleului celular și este una cu aceasta. Astfel, cavitățile reticulului endoplasmatic se deschid în cavitatea intermembranară a membranei nucleare. Membranele EPS asigură transportul activ al unui număr de elemente împotriva unui gradient de concentrație. Filamentele care formează reticulul endoplasmatic au un diametru de 0,05-0,1 µm (uneori până la 0,3 µm), grosimea membranelor cu două straturi care formează peretele tubilor este de aproximativ 50 angstromi (5 nm, 0,005 µm). Aceste structuri conțin fosfolipide nesaturate, precum și unele colesterol și sfingolipide. Conțin și proteine.
Tubulii, al căror diametru variază de la 0,1-0,3 µm, sunt umpluți cu conținut omogen. Funcția lor este implementarea comunicării între conținutul veziculelor EPS, mediul extern și nucleul celular.
Reticulul endoplasmatic nu este o structură stabilă și este supus unor modificări frecvente.
Există două tipuri de EPR:
- reticul endoplasmatic granular
- reticul endoplasmatic agranular (neted).
Pe suprafața reticulului endoplasmatic granular există un număr mare de ribozomi, care sunt absenți pe suprafața ER agranulară.
Reticulul endoplasmatic granular și agranular îndeplinesc diferite funcții în celulă.
Funcțiile reticulului endoplasmatic
Cu participarea reticulului endoplasmatic, au loc translația și transportul proteinelor, sinteza și transportul lipidelor și steroizilor. EPS se caracterizează și prin acumularea de produse de sinteză. Reticulul endoplasmatic este, de asemenea, implicat în crearea unei noi membrane nucleare (de exemplu, după mitoză). Reticulul endoplasmatic conține o cantitate intracelulară de calciu, care este, în special, un mediator al contracției celulelor musculare. În celulele fibrelor musculare există o formă specială a reticulului endoplasmatic - reticulul sarcoplasmic.
Funcțiile reticulului endoplasmatic agranular
Reticulul endoplasmatic agranular este implicat în multe procese metabolice. Enzimele reticulului endoplasmatic agranular sunt implicate în sinteza diferitelor lipide și fosfolipide, acizi grași și steroizi. De asemenea, reticulul endoplasmatic agranular joaca un rol important in metabolismul carbohidratilor, dezinfectarea celulelor si depozitarea calciului. În special, în legătură cu aceasta, reticulul endoplasmatic agranular predomină în celulele glandelor suprarenale și ficatului.
Sinteza hormonilor
Hormonii care se formează în EPS agranular includ, de exemplu, hormoni sexuali de vertebrate și hormoni steroizi suprarenali. Celulele testiculare și ovariene responsabile de sinteza hormonală conțin cantități mari de reticul endoplasmatic agranular.
Acumularea și conversia carbohidraților
Carbohidrații din organism sunt stocați în ficat sub formă de glicogen. Glicoliza transformă glicogenul din ficat în glucoză, care este un proces critic în menținerea nivelului de glucoză din sânge. Una dintre enzimele EPS agranulare desprinde o grupare fosfo din primul produs al glicolizei, glucoza-6-fosfat, permițând astfel glucozei să părăsească celula și să crească nivelul zahărului din sânge.
Neutralizarea otrăvurilor
Reticulul endoplasmatic neted al celulelor hepatice Participarea activăîn neutralizarea tot felul de otrăvuri. Enzimele netede ER ataseaza moleculele de substante active intalnite, care pot fi astfel dizolvate mai rapid. În cazul aportului continuu de otrăvuri, droguri sau alcool se formează o cantitate mai mare de EPR agranular, care crește doza de substanță activă necesară pentru a obține același efect.
Reticulul sarcoplasmic
O formă specială a reticulului endoplasmatic agranular, reticulul sarcoplasmatic, formează ER în celulele musculare, în care ionii de calciu sunt pompați activ din citoplasmă în cavitatea ER împotriva gradientului de concentrație în starea neexcitată a celulei și eliberați în citoplasmă. pentru a iniția contracția. Concentrația ionilor de calciu în EPS poate ajunge la 10 −3 moli, în timp ce în citosol este de aproximativ 10 −7 moli (în repaus). Astfel, membrana reticulului sarcoplasmatic asigură transport activ împotriva gradienților de concentrație de ordin înalt. Iar aportul și eliberarea ionilor de calciu în EPS este într-o relație subtilă cu condițiile fiziologice.
Concentrația ionilor de calciu în citosol afectează multe procese intracelulare și intercelulare, cum ar fi: activarea sau inhibarea enzimelor, expresia genelor, plasticitatea sinaptică a neuronilor, contracțiile celulelor musculare, eliberarea de anticorpi din celulele sistemului imunitar.
Funcțiile reticulului endoplasmatic granular
Reticulul endoplasmatic granular are două funcții: sinteza proteinelor și producerea membranei.
Sinteza proteinei
Proteinele produse de celulă sunt sintetizate pe suprafața ribozomilor, care pot fi atașați la suprafața RE. Lanțurile polipeptidice rezultate sunt plasate în cavitatea reticulului endoplasmatic granular (unde cad și lanțurile polipeptidice sintetizate în citosol), unde sunt ulterior tăiate și pliate în mod corect. Astfel, secvențele liniare de aminoacizi se obțin după translocarea în reticulul endoplasmatic a structurii tridimensionale necesare, după care sunt retransferate în citosol.
Sinteza membranei
Ribozomii atașați la suprafața ER granulară produc proteine, care, împreună cu producția de fosfolipide, printre altele, extinde suprafața membranei proprii a ER, care trimite fragmente de membrană către alte părți ale sistemului membranar prin vezicule de transport.
Vezi si
- Reticulonii sunt proteine ale reticulului endoplasmatic.
Fundația Wikimedia. 2010 .
Închiderea se referă la conexiune strânsă(zona de inchidere - zonu-la occuludens). Această conexiune implică proteine integrale speciale situate pe suprafața celulelor învecinate, formând o aparență de rețea de plasă (Fig. 4.6).
Această rețea celulară înconjoară întregul perimetru al celulei sub forma unei centuri, conectându-se cu aceeași rețea pe suprafața celulelor învecinate. Această zonă este impenetrabilă pentru macromolecule și ioni și, prin urmare, se închide, delimitează golurile intercelulare (și, împreună cu acestea, mediul intern al organismului) de mediul extern. Acest tip de conexiuni este tipic pentru celulele epiteliului cu un singur strat și endoteliul unor vase.
Conexiunile de aderență sau ancorare includ bandă adezivă(breaua de aderenta) si desmozom. Comun acestui grup de compuși este faptul că elementele fibrilare ale citoscheletului (filamente de actină,
Orez. 4.6. Conexiune strânsă (zona de închidere):
dar- localizarea joncțiunii strânse (placă de inserție) pe celulele epiteliului intestinal; b - diagrama tridimensională a unei zone dense de îmbinare. 1 - microvilozități
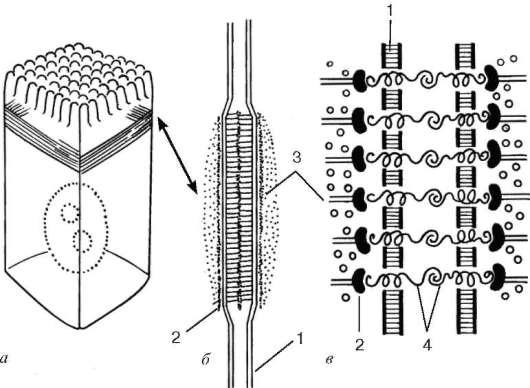
Orez. 4.7. Banda adeziva (banda de aderenta):
dar- localizarea acestuia în celulă; b- vedere în sectiune; în- schema de organizare moleculară. 1 - plasmalema; 2 - strat de proteine de adeziune; 3 - microfilamente de actină; 4 - glicoproteine de legare
filamentele intermediare și spectrina) și se leagă de membrane la joncțiunea celulelor învecinate.
banda adeziva, sau centură aglomerată (zonula adherens),- o formațiune pereche sub formă de panglici, fiecare din care înconjoară părțile apicale ale celulelor învecinate și asigură aderența lor între ele în această zonă (Fig. 4.7). Aici, celulele sunt conectate între ele prin glicoproteine integrale, la care se învecinează un strat de proteine membranare, inclusiv vinculină, proteina caracteristică, din partea citoplasmei ambelor celule. Un mănunchi de microfilamente de actină se apropie de acest strat și se leagă de el. Interacțiunea microfilamentelor de actină cu proteinele care leagă actina din multe celule învecinate poate duce la o modificare a reliefului întregului strat epitelial.
Conexiunile adezive pot include contact focal, caracteristice fibroblastelor. În acest caz, celula nu se conectează cu o celulă vecină, ci cu elemente ale substratului extracelular. Microfilamentele de actină participă și ele la formarea contactului focal. Joncțiunile intercelulare adezive includ desmozomi(Fig. 4.8).
desmozom, sau punct de adeziune (macula adherens). Acestea sunt structuri pereche, care sunt o zonă mică sau un punct cu un diametru de aproximativ 0,5 µm. Din partea citoplasmei, un strat de proteine, care include desmoplakine, este adiacent membranei plasmatice. Mănunchiuri de filamente intermediare sunt introduse în acest strat din partea laterală a citoplasmei. Pe partea exterioară a plasmolemei celulelor vecine din regiunea desmozomilor sunt conectate
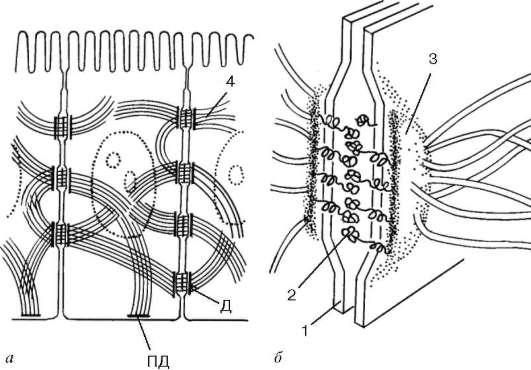
Orez. 4.8. Desmosom:
dar- amplasarea in cusca; b- schema ultrastructurii. 1 - plasmalema; 2 - stratul desmo-gley; 3 - strat de desmoplakin; 4 - filamente intermediare. D - desmozom; PD - hemidesmozom
cu ajutorul proteinelor transmembranare – desmogleine. De exemplu, fiecare celulă a epidermei pielii poate avea până la câteva sute de desmozomi.
Rolul funcțional al desmozomilor este în principal în legătura mecanică dintre celule. Desmozomii leagă celulele între ele în diferite epitelii, în mușchii cardiaci și netezi. Hemidesmozomi leagă celulele epiteliale de membrana bazală.
Conexiunile de comunicare în celulele animale sunt reprezentate de joncțiuni gap și sinapse(Fig. 4.9).
conexiune întreruptă, sau nexus (nexus), reprezintă o regiune cu lungimea de 0,5-3 microni, unde membranele plasmatice sunt separate printr-un interval de 2-3 nm (vezi Fig. 4.9). Din partea citoplasmei, nu se găsesc structuri speciale legate de membrană în această zonă, dar în structura plasmolemelor celulelor învecinate, complexe proteice speciale (conexiuni) sunt situate unul față de celălalt, care formează canale de la o celulă la alta. . Acest tip de conexiune se găsește în toate grupele de țesuturi.
Rolul funcțional al joncțiunii gap este transferul de ioni și molecule mici (greutate moleculară 2 × 103) de la celulă la celulă. Deci, în mușchiul inimii, excitația, care se bazează pe procesul de modificare a permeabilității ionilor, este transmisă de la celulă la celulă prin nexusuri.
Conexiuni sinaptice sau sinapse (synapsis). Acest tip de conexiuni este caracteristic țesutului nervos și apare în zone specializate de contact atât între doi neuroni, cât și între un neuron și un alt element care face parte dintr-un receptor sau efector (de exemplu, sinapsele neuromusculare, neuroepiteliale).
Sinapsele sunt zone de contact între două celule specializate pentru transmiterea într-un singur sens a excitației sau inhibiției de la un element la altul (vezi capitolul 10).
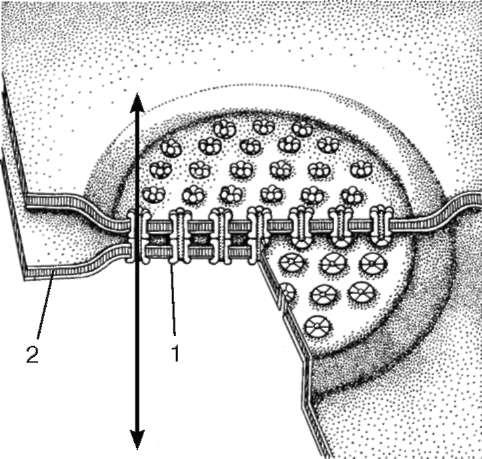
Orez. 4.9. Conexiune cu fante (comunicare):
1 - conexon; 2 - plasmalema
sistemul vacuolar
Reticulul endoplasmatic
Reticulul endoplasmatic (reticulul endoplasmatic) a fost descoperit de K. R. Porter în 1945. Această componentă a sistemului vacuolar celular este o colecție de vacuole, saci cu membrană plate sau formațiuni tubulare care creează o rețea de membrană tridimensională. Rețeaua include granulară și agranulară secțiuni care pot fi intercalate.
Reticulum endoplasmatic granular (reticulum endoplasmicum granulosum) pe secțiuni ultrasubțiri este reprezentată de membrane închise, care formează pungi turtite, cisterne, tuburi pe secțiuni.
Diametrul cisternelor variază semnificativ și, în funcție de activitatea funcțională a celulei, variază de la 20 nm la câțiva micrometri. O caracteristică distinctivă a membranelor reticulului endoplasmatic granular este aceea că sunt acoperite cu numeroși ribozomi din partea hialoplasmei (Fig. 4.10).
Reticulul endoplasmatic granular are o structură diferită. Pentru celulele nespecializate sau pentru celulele cu activitate metabolică scăzută este caracteristică prezența cisternelor rare și împrăștiate. Dacă există acumulări locale ale reticulului endoplasmatic granular, atunci aceasta indică o sinteză activă a proteinelor secretoare. Deci, în celulele hepatice și unele celule nervoase, reticulul endoplasmatic granular este asamblat în zone separate. În celulele pancreasului, reticulul endoplasmatic granular sub formă de cisterne membranare strâns strâns unul lângă celălalt ocupă zonele bazale și perinucleare ale celulei. Ribozomii asociați cu membranele reticulului endoplasmatic sunt implicați în sinteza proteinelor care sunt excretate dintr-o anumită celulă (proteinele „exportate”). În plus, reticulul endoplasmatic granular este implicat în sinteza proteinelor - enzime necesare organizării metabolismului intracelular și, de asemenea, utilizate pentru digestia intracelulară.
Proteinele care se acumulează în cavitățile reticulului endoplasmatic pot, ocolind hialoplasma, să fie transportate în vacuolele complexului Golgi, unde sunt modificate și fac parte fie din lizozomi, fie din granulele secretoare, al căror conținut rămâne izolat de hialoplasmă prin membrana. În interiorul tubilor sau vacuolelor endoplasmatice granulare
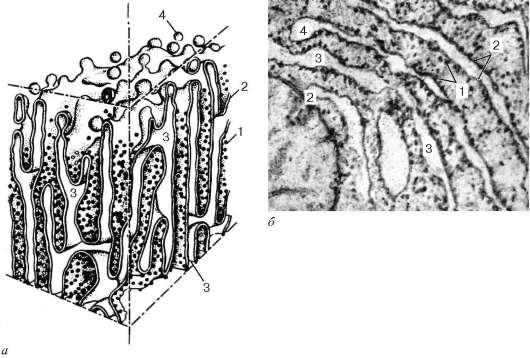
Orez. 4.10. Structura reticulului endoplasmatic granular: dar - sistem; b- micrografie electronică a unei secțiuni a unei celule epiteliale hepatice. 1 - ribozomi; 2 - farfurii; 3 - cavitățile interne ale rezervoarelor; 4 - desprinderea veziculelor membranare lipsite de ribozomi
de rețea, proteinele sunt modificate, de exemplu, prin legarea lor de zaharuri (glucozilare primară).
În reticulul endoplasmatic granular, pe ribozomii săi, se sintetizează proteine integrale membranare, care sunt înglobate în grosimea membranei. Aici, din partea hialoplasmei, are loc sinteza lipidelor și încorporarea lor în membrană. Ca rezultat al acestor două procese, membranele reticulului endoplasmatic și alte componente ale sistemului vacuolar al celulei cresc.
Deci, rolul reticulului endoplasmatic granular constă în sinteza proteinelor exportate pe ribozomii săi, în izolarea acestora de conținutul hialoplasmei din interiorul cavităților membranei, în transportul acestor proteine în alte părți ale celulei, în substanța chimică. modificarea unor astfel de proteine și în condensarea lor locală, precum și în sinteza componentelor structurale ale membranelor celulare.
reticul endoplasmatic agranular (neted) (reticul endoplasmaticum nongranulosum) reprezentata si de membrane care formeaza mici vacuole, tuburi, tubuli, care se pot ramifica, se contopesc intre ele. Spre deosebire de ER granular, nu există ribozomi pe membranele ER netede. Diametrul vacuolelor și tubilor reticulului endoplasmatic neted este de obicei de aproximativ 50-100 nm. Reticulul endoplasmatic neted apare și se dezvoltă pe baza reticulului endoplasmatic granular. În unele zone ale reticulului endoplasmatic granular se formează noi zone membranare lipoproteice, lipsite de ribozomi. Aceste zone pot crește, se pot separa de membranele granulare și pot funcționa ca un sistem vacuolar independent.
Activitatea reticulului endoplasmatic neted este asociată cu metabolismul lipidelor și a unor polizaharide intracelulare. Reticulul endoplasmatic neted este implicat în etapele finale ale sintezei lipidelor. Este foarte dezvoltat în celulele secretoare de steroizi, de exemplu, în celulele endocrine ale cortexului suprarenal, în celulele epiteliale ale tubilor seminiferi contorți.
Legătura topografică strânsă a reticulului endoplasmatic neted cu depozitele de glicogen (polizaharidă intracelulară de rezervă a animalelor) în hialoplasma diferitelor celule (celule hepatice, fibre musculare) indică posibila sa participare la metabolismul carbohidraților.
În fibrele musculare striate, reticulul endoplasmatic neted este capabil să depună ioni de calciu necesari pentru funcționarea țesutului muscular (vezi capitolul 9).
Rolul reticulului endoplasmatic neted în dezactivarea diferitelor substanțe dăunătoare organismului datorită oxidării lor cu ajutorul unui număr de enzime speciale este foarte important. Mai ales clar se manifestă în celulele ficatului. Deci, cu unele otrăviri, în celulele hepatice apar zone acidofile (nu conțin ARN), complet umplute cu un reticul endoplasmatic neted.
Complexul Golgi
Complexul Golgi (complex lamelar) a fost descoperit în 1898 de K. Golgi. Autorul, folosind proprietățile de legare ale metalelor grele (osmiu sau argint) cu structurile celulare, a dezvăluit formațiuni nete în celulele nervoase, pe care le-a numit aparatul rețelei interne. (aparatul reticular intern). Mai târziu s-a numit aparat, sau Complexul Golgi (complexul Golgiensis). Structuri similare au fost apoi descrise în toate celulele eucariote.
Când este privit la microscop electronic, complexul Golgi este reprezentat de structuri membranare asamblate împreună în zone mici.
Se numește o zonă separată de acumulare a acestor membrane dictiozom (stiva Golgi).Într-o celulă pot exista mai multe astfel de zone. Aproape unul de celălalt (la o distanță de 20-25 nm) sunt 5-10 plate cisterne,între care se află straturi subțiri de hialoplasmă. Fiecare rezervor are o grosime variabilă: în centrul membranelor sale pot fi reunite (până la 25 nm), iar la periferie pot avea prelungiri - fiole, a căror lățime nu este constantă. Pe lângă cisternele plate distanțate dens, în zona complexului Golgi sunt observate multe bule mici. (veziculă), care se găsesc mai ales în zonele sale periferice. Uneori sunt dantelate din prelungiri ampulare la marginile cisternelor plate. În zona dictiozomului, cea proximală (cis)și distal (trans) suprafete. În celulele secretoare, complexul Golgi este de obicei polarizat: suprafața sa proximală este orientată spre nucleu, în timp ce cea distală este orientată spre suprafața celulei.
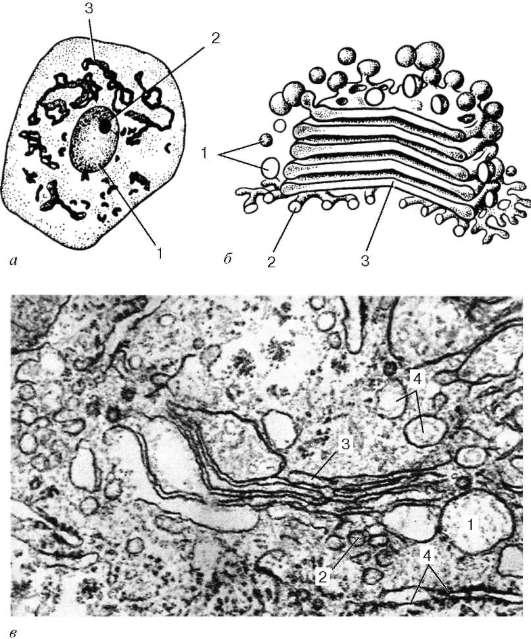
Orez. 4.11. Complexul Golgi:
dar - celula nervoasa măduva spinării, impregnare cu argint după metoda Golgi: 1 - miez; 2 - nucleol; 3 - complexul Golgi; b- schema structurii ultramicroscopice (reconstructie tridimensionala); în- complex Golgi pe o secțiune ultrasubțire (celula hepatică): 1 - vezicule; 2 - tubuli; 3 - pungi turtite (rezervoare); 4 - fragmente ale reticulului endoplasmatic granular
În celule, dictiozomii individuali pot fi conectați între ei printr-un sistem de vezicule și cisterne adiacente suprafeței distale, astfel încât se formează o rețea tridimensională liberă, care este detectată în microscoapele luminoase și electronice („rețeaua trans” de complexul Golgi).
Complexul Golgi este implicat în segregarea și acumularea produselor sintetizate în reticulul endoplasmatic, în rearanjarea chimică, maturarea acestora; în rezervoarele sale, polizaharidele sunt sintetizate și complexate cu proteine, ceea ce duce la formarea peptidoglicanilor. Cu ajutorul complexului Golgi, se realizează procesul de îndepărtare a secretelor gata făcute în afara celulei secretoare. În plus, complexul
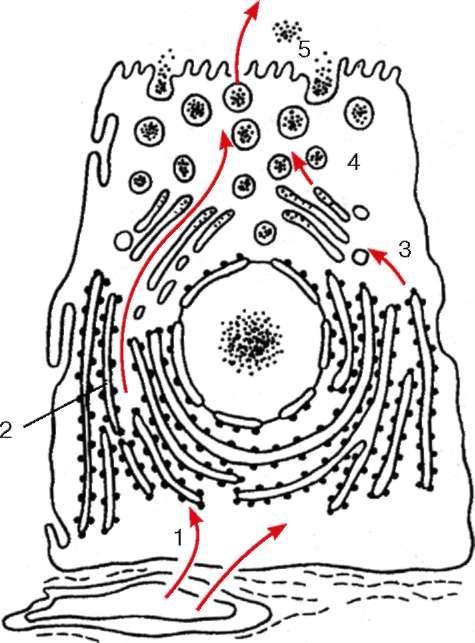
Orez. 4.12. Participarea structurilor celulare la secreția proteică (schemă): 1 - alimentarea cu aminoacizi din hemocapilar către ribozomii reticulului endoplasmatic granular; 2 - sinteza si segregarea proteinelor; 3 - tranziția proteinelor în vacuolele complexului Golgi; 4 - clivaj din complexul Golgi al veziculelor cu produse secretoare; 5 - eliberarea secretului din celulă
Golgi asigură formarea lizozomilor. Membranele complexului sunt formate prin scindarea unor mici vacuole din reticulul endoplasmatic granular. Aceste vacuole intră în complexul Golgi proximal, unde se contopesc cu membranele acestuia. În consecință, noi porțiuni de membrane și produse sintetizate în reticulul endoplasmatic granular intră în complexul Golgi. În cisternele membranare ale complexului Golgi apar modificări secundare în structura proteinelor sintetizate în reticulul endoplasmatic granular. Aceste modificări (modificări) sunt asociate cu rearanjarea lanțurilor de oligozaharide ale glicoproteinelor sintetizate. În interiorul cavităților complexului Golgi, cu ajutorul diferitelor enzime (trans-glucozidaze), proteinele lizozomale și proteinele de secreție sunt modificate în diferite moduri: au loc înlocuirea secvențială și creșterea lanțurilor de oligozaharide. Proteinele modificatoare trec de la cisterna suprafeței cis proximale la cisternele suprafeței distale prin transferul în cursă de releu a vacuolelor mici care conțin proteina transportată.
În cisterne distale (trans) La suprafață, proteinele sunt sortate: pe suprafețele interioare ale membranelor rezervoarelor există receptori care recunosc fie proteine secretoare, fie proteine care alcătuiesc lizozomii (hidrolaze). Ca urmare, două tipuri de vacuole mici se desprind din cisternele suprafeței distale a dictiozomilor: a) care conțin hidrolaze - lizozomi (primare); b) proteine secretoare.
Funcția secretorie a complexului Golgi este aceea că proteina sintetizată pe ribozomi, care se acumulează în interiorul cisternelor reticulului endoplasmatic, este transportată mai departe în vacuolele complexului Golgi (Fig. 4.12).
Apoi proteina acumulată se poate condensa, formând produse proteice secretoare (cum se observă, de exemplu, în pancreas, glande mamare și alte glande). Din prelungiri ampulare ale rezervoarelor complexului
sa Golgi a desprins veziculele care conțin aceste proteine. În viitor, ele pot fuziona între ele și endozomii și pot crește în dimensiune, formându-se granule secretoare. După aceea, granulele secretoare încep să se deplaseze spre suprafața celulei, vin în contact cu membrana plasmatică, cu care se contopesc propriile membrane și astfel conținutul granulelor se află în afara celulei. Din punct de vedere morfologic, acest proces se numește extrudare (ejecție, exocitoză) și seamănă cu pinocitoza doar cu succesiunea inversă a etapelor.
Trebuie remarcat faptul că din momentul formării și până la excreția din celule, produsele secretate sunt separate printr-o membrană de hialoplasmă. Prin urmare, membranele complexului Golgi îndeplinesc un rol segregator în formarea secrețiilor celulare. În vacuolele complexului Golgi, uneori are loc o acumulare de molecule de lipide resintetizate și formarea de proteine complexe - lipoproteine, care pot fi transportate de vacuole în afara celulei. Vacuolele complexului Golgi dau naștere la lizozomi.
Lizozomi
Lizozomi (lizozomi)- aceasta este o clasă diversă de vacuole cu dimensiunea de 0,2-0,4 microni, limitate de o singură membrană. trăsătură caracteristică lizozomii este prezența în ei a enzimelor hidrolitice - hidrolaze (proteinaze, nucleaze, fosfataze, lipaze etc.), care descompun diversi biopolimeri la o valoare a pH-ului acid. Lizozomii au fost descoperiți în 1949 de de Duve.
Pe lângă lizozomii înșiși (primari), se disting autofagolizozomii sau heterolizozomii (lizozomii secundari) și telolizozomii (corpii reziduali) (Fig. 4.13).
Diversitatea morfologiei lizozomilor se explică prin faptul că aceste particule sunt implicate în procesele de digestie intracelulară, formând vacuole digestive complexe de origine atât exogenă (extracelulară), cât și endogenă (intracelulară).
Lizozomi (primari) sunt mici vezicule membranare de aproximativ 0,2-0,5 microni, umplute cu o substanță fără structură care conține hidrolaze, inclusiv fosfatază acidă activă, care este o enzimă marker pentru lizozomi. Aceste vezicule mici sunt practic foarte greu de distins de veziculele mici de la periferia zonei Golgi, care conțin și fosfatază acidă. Locul sintezei sale este reticulul endoplasmatic granular. Apoi, această enzimă apare în cisternele suprafeței proximale a dictiozomului și apoi în vezicule mici de-a lungul periferiei dictiozomului și, în final, în lizozomi. Astfel, întreaga cale de formare a lizozomului este foarte asemănătoare cu formarea granulelor secretoare (zimogene) în celulele pancreatice, cu excepția ultimei etape.
heterofagolizozomi (lizozomi secundari) sau vacuole digestive intracelulare, sunt formate prin fuziunea lizozomilor cu vacuole fagocitare sau pinocitare. Dacă are loc fuziunea lizozomilor
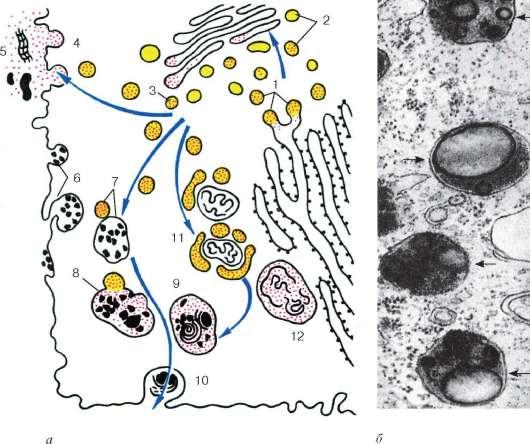
Orez. 4.13. Structura lizozomilor:
dar - schema de participare a structurilor celulare la formarea lizozomilor și la digestia intracelulară: 1 - formarea de vezicule mici care conțin enzime hidrolitice din reticulul endoplasmatic granular; 2 - transferul enzimelor în complexul Golgi; 3 - formarea lizozomilor primari; 4 - izolarea și utilizarea (5) hidrolazelor în clivaj extracelular; 6 - vezicule endocitare; 7 - fuziunea lizozomilor primari și veziculelor endocitare; 8 - formarea lizozomilor secundari; 9 - telolizozomi; 10 - excreția corpurilor reziduale; 11 - fuziunea lizozomilor primari cu structurile celulare în colaps; 12 - autofagolizozom; b - Micrografie electronică a unei secțiuni de heterofagolizozomi (indicată prin săgeți)
cu organele modificate ale celulei în sine, atunci se numește o astfel de structură autofagolizozom.În acest caz, enzimele lizozomilor au acces la substraturi, pe care încep să le descompună. Substanțele care au intrat în compoziția hetero sau autofagolizozomilor (lizozomi secundari) sunt scindate de hidrolaze la monomeri, care sunt transportați prin membrana lizozomului la hialoplasmă, unde sunt reutilizate, adică sunt incluse în diferite procese metabolice.
Cu toate acestea, scindarea și digestia macromoleculelor de către lizozom pot să nu se finalizeze într-un număr de celule. În acest caz, în vacuolele lizozomilor se acumulează
alimente nedigerate. Acest organel se numește corp lizozom, sau corp rezidual (corpusculum residuale). Corpurile reziduale conțin mai puține enzime hidrolitice, conțin compactarea conținutului, restructurarea acestuia. Adesea în corpurile reziduale se observă structurarea secundară a lipidelor nedigerabile, care formează structuri stratificate. Acolo se depun si pigmenti. De exemplu, la om, în timpul îmbătrânirii, în celulele creierului, ficatului și fibrelor musculare din telolizozomi, există o depunere de „pigment de îmbătrânire” - lipofuscină.
Cu participarea lizozomilor (autofagolizozomi), poate avea loc modificarea produselor care sunt sintetizate de celula însăși. Astfel, cu ajutorul enzimelor lizozomale, tiroglobulina este hidrolizată în celulele glandei tiroide, ceea ce duce la formarea hormonilor tiroidieni, care sunt apoi excretați în sânge prin exocitoză.
ÎN autofagolizozomi Se găsesc fragmente sau chiar structuri citoplasmatice întregi, precum mitocondrii, elemente ale reticulului endoplasmatic, ribozomi, granule de glicogen și altele, ceea ce este dovada rolului lor decisiv în procesele de digestie intracelulară.
Semnificația funcțională a autofagocitozei este încă neclară. Există o presupunere că acest proces este asociat cu selecția și distrugerea componentelor celulare alterate, deteriorate. În acest caz, lizozomii joacă rolul de „curățători” intracelulari care îndepărtează structurile defecte. Interesant, în condiții normale, numărul de autofagolizozomi crește sub stres metabolic, de exemplu, în timpul inducerii hormonale a activității celulelor hepatice. Numărul de autofagolizozomi crește semnificativ cu diferite leziuni celulare; în acest caz, zone întregi din interiorul celulelor pot suferi autofagocitoză.
O creștere a numărului de autofagolizozomi în celule în timpul proceselor patologice este un fenomen comun.
Peroxizomii
Peroxizomi (peroxisomae)în celulele țesutului uman, acestea sunt corpuri mici (0,3-1,5 μm în dimensiune) de formă ovală, limitate de o membrană, care conțin o matrice granulară, în centrul căreia sunt adesea structuri cristaline formate din fibrile și tuburi (miez). vizibil. Peroxizomii sunt caracteristici în special celulelor hepatice și renale. În fracția peroxizomală se găsesc enzime de oxidare a aminoacizilor, în timpul cărora se formează peroxid de hidrogen și se detectează și enzima catalaza, care o distruge. Catalaza peroxizomului joacă un rol protector important, deoarece H 2 O 2 este o substanță toxică pentru celulă.
Astfel, organele celulare cu o singură membrană care alcătuiesc sistemul vacuolar asigură sinteza și transportul biopolimerilor intracelulari, produse de secreție excretate din celulă, care este însoțită de biosinteza tuturor membranelor acestui sistem. Lizozomii și peroxizomii sunt implicați în degradarea substraturilor celulare exogene și endogene.
mitocondriile
Mitocondriile (mitocondriale)- sistemul energetic al celulei, organitele sintezei ATP. Funcția lor principală este asociată cu oxidarea compușilor organici și utilizarea energiei eliberate în timpul dezintegrarii acestor compuși pentru sinteza moleculelor de ATP. Pe baza acestui fapt, mitocondriile sunt adesea numite stațiile energetice ale celulei sau organelele respirației celulare.
Termenul „mitocondrii” a fost introdus de Benda în 1897 pentru a desemna structurile granulare și filamentoase din citoplasma diferitelor celule. Mitocondriile pot fi observate în celulele vii, deoarece au o densitate destul de mare. Forma și dimensiunea mitocondriilor celulelor animale sunt variate, dar în medie grosimea lor este de aproximativ 0,5 microni, iar lungimea lor este de la 1 la 10 microni. Calculele arată că numărul lor în celule variază foarte mult - de la elemente individuale la sute. Deci, în celula hepatică, ele reprezintă mai mult de 20% din volumul total al citoplasmei și conțin aproximativ 30-35% din cantitatea totală de proteine din celulă. Suprafața tuturor mitocondriilor celulei hepatice este de 4-5 ori mai mare decât suprafața membranei plasmatice.
În multe cazuri, mitocondriile individuale pot avea dimensiuni gigantice și reprezintă o rețea extinsă - reticulul mitocondrial. Deci, de exemplu, în muschii scheletici reticulul mitocondrial este reprezentat de multe catene mitocondriale ramificate si gigantice. Mitocondriile ramificate gigantice se găsesc în celulele nefronilor proximali etc.
Mitocondriile se acumulează de obicei în apropierea acelor părți ale citoplasmei unde este nevoie de ATP. Deci, în mușchiul inimii, mitocondriile sunt situate în apropierea miofibrilelor. La spermatozoizi, mitocondriile formează o carcasă spirală în jurul axei flagelului etc. O creștere a numărului de mitocondrii din celule are loc prin diviziunea sau înmugurirea mitocondriilor originale.
Mitocondriile sunt limitate de două membrane de aproximativ 7 nm grosime (Fig. 4.14).
Membrana mitocondrială exterioară (membrana mitocondrială externă) le separă de hialoplasmă. De obicei are contururi uniforme și este închisă, astfel încât să fie o pungă cu membrană. Membrana exterioară este separată de cea interioară printr-un spațiu intermembranar de aproximativ 10–20 nm lățime. Membrana mitocondrială internă (membrana mitocondrialis interna) limitează conținutul intern real al mitocondriilor, acesta matrice (matrice mitocondrială). O trăsătură caracteristică a membranelor interioare ale mitocondriilor este capacitatea lor de a forma numeroase proeminențe în interiorul mitocondriilor. Astfel de proeminențe arată cel mai adesea ca niște creste plate sau crist.
Matricea mitocondrială are o structură cu granulație fină (vezi Fig. 4.14, b) uneori dezvăluie filamente subțiri (aproximativ 2-3 nm grosime) și granule de aproximativ 15-20 nm în dimensiune. Catenele matricei mitocondriale sunt molecule de ADN, iar granulele mici sunt ribozomi mitocondriali.
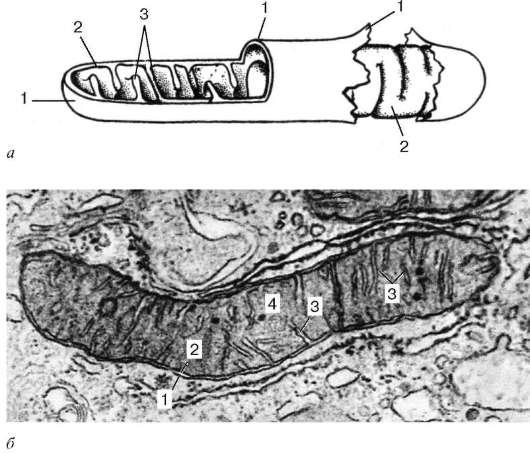
Orez. 4.14. Structura ultramicroscopică a mitocondriilor:
dar- schema; b- micrografie electronică a unei secțiuni a mitocondriei unei celule hepatice. 1 - membrana mitocondrială exterioară; 2 - membrana mitocondriala interioara; 3 - cristae; 4 - matricea mitocondrială
Funcția principală a mitocondriilor este sinteza ATP, care are loc ca urmare a proceselor de oxidare a substraturilor organice și de fosforilare a adenozindifosfatului (ADP).
Stadiile inițiale ale acestor procese complexe au loc în hialoplasmă. Aici, oxidarea primară a substraturilor (de exemplu, zaharuri) la acid piruvic (piruvat) are loc cu sinteza simultană a unei cantități mici de ATP. Aceste procese au loc în absența oxigenului (oxidare anaerobă, glicoliză). Toate etapele ulterioare ale producției de energie - oxidarea aerobă și sinteza majorității ATP - sunt efectuate cu consum de oxigen și sunt localizate în interiorul mitocondriilor. În acest caz, oxidarea ulterioară a piruvatului și a altor substraturi ale metabolismului energetic are loc odată cu eliberarea de CO 2 și transferul de protoni către acceptorii lor. Aceste reacții sunt efectuate cu ajutorul unui număr de enzime din așa-numitul ciclu al acidului tricarboxilic, care sunt localizate în matricea mitocondrială.













