Cine are reticulul endoplasmatic. Organele de importanță generală. Reticulul endoplasmatic
1. Cărui grup de organite aparțin lizozomii, reticulul endoplasmatic si aparatul Golgi?
Ele aparțin organelelor cu o singură membrană.
2. Care este structura și funcția reticulului endoplasmatic? Cum este EPS aspru diferit de neted?
Reticulul endoplasmatic (EPS) sau reticulul endoplasmatic (ER), este un sistem de canale și cavități înconjurate de o membrană și care pătrund în hialoplasma celulei. Distingeți între EPS aspru și neted. Reticulul endoplasmatic dur conține mulți ribozomi. Aici sunt sintetizate proteinele care sunt excretate în afara celulei. Proteinele membranare sunt de asemenea sintetizate pe ribozomii ER brut. Sinteza carbohidraților și a lipidelor are loc pe suprafața EPS neted. În plus, ionii de calciu, care sunt regulatori importanți ai funcțiilor celulare și ale corpului în ansamblu, se acumulează în ER neted. Substanțele care se formează pe membranele EPS se acumulează în interiorul cavităților rețelei și sunt transformate. De exemplu, proteinele capătă structura lor caracteristică secundară, terțiară sau cuaternară. EPS dur este mai bine dezvoltat în acele celule care sintetizează un numar mare de proteine (de exemplu, în celulele glandelor salivare și pancreasului, care realizează sinteza enzimelor digestive; în celulele pancreasului și glandei pituitare). Smooth ER este bine dezvoltat în celulele care sintetizează, de exemplu, polizaharide și lipide (celule ale glandelor suprarenale și ale gonadelor care produc hormoni steroizi; celule hepatice care sintetizează glicogen etc.)
3. Cum funcționează complexul Golgi? Ce funcții îndeplinește?
Complexul (aparatul) Golgi este un sistem de structuri membranare intracelulare: rezervoare și vezicule, în care se acumulează substanțe sintetizate pe membranele EPS. Substanțele sunt livrate în complexul Golgi în vezicule membranoase care se desprind de reticulul endoplasmatic și se unesc cu cisternele complexului Golgi. Aici, aceste substanțe suferă diferite transformări biochimice și apoi sunt din nou împachetate în vezicule membranare, iar cele mai multe dintre ele sunt transportate în membrana citoplasmatică. Membrana veziculelor fuzionează cu membrana citoplasmatică, iar conținutul este expulzat în afara celulei prin exocitoză. În complexul Golgi celule vegetale se sintetizează polizaharidele din peretele celular (cochilii). O altă funcție importantă a complexului Golgi este formarea lizozomilor.
4. Majoritatea complexe mari Golgi (până la 10 microni) se găsesc în celulele glandelor endocrine. Ce crezi, cu ce este legat?
Glandele endocrine produc hormoni care sunt necesari proceselor vitale ale organismului. Aceeași formare a unui număr de hormoni și maturarea lor se realizează în cavitățile complexului Golgi.
5. Ce este comun în structura și funcțiile reticulului endoplasmatic și complexului Golgi? Care este diferența?
Similitudine: sunt organite cu o singură membrană și reprezintă un sistem de cavități și cisterne. Diferențe: în EPS are loc formarea de substanțe, care apoi intră în complexul Golgi și suferă transformări biochimice (se coace). De asemenea, complexul Golgi este implicat în formarea lizozomilor.
6. Ce sunt lizozomii? Cum sunt formate? Ce funcții îndeplinesc?
Lizozomii sunt mici vezicule membranoase care se împletesc din cisternele complexului Golgi și conțin un set de enzime digestive. Enzimele lizozomilor sunt capabile să descompună proteinele, carbohidrații, lipidele, acizii nucleici. Pe lângă digerarea substanțelor care intră în celulă din exterior, lizozomii participă la descompunerea componentelor interne ale celulei (molecule și organite întregi), deteriorate sau învechite. Acest proces se numește autofagie.
7. Sugerați de ce enzimele din lizozom nu își scindează propria membrană. Care sunt consecințele pentru celula care poate duce la ruperea membranelor lizozomului?
Lizozomii conțin un set extins de enzime hidrolitice capabile să descompună aproape toate tipurile de macromolecule biologice. În mod normal, aceste enzime nu acționează asupra componentelor propriilor celule, deoarece sunt separate de o membrană lizozomală. Când membrana lizozomului se rupe, enzimele din aceasta intră în celulă și încep să absoarbă și să digere conținutul său intern, ceea ce duce la moartea celulei.
8. S-a stabilit că anumite oligo- sau polizaharide sunt „atașate” de moleculele multor substanțe pentru a fi îndepărtate din celulă din complexul Golgi, iar diferite componente glucide sunt atașate la diferite substanțe. În această formă modificată, substanțele sunt excretate în mediul extracelular. De ce crezi că este nevoie de asta?
maturizarea are loc în complexul Golgi diverse substante, care sunt apoi trimise la diferite organite ale celulei sau îndepărtate din aceasta. Un astfel de proces treptat trebuie cumva controlat. Prin urmare, substanțele maturizate sunt marcate cu oligo- și polizaharide speciale, care joacă rolul unui fel de marcă de calitate. Cu ajutorul acestora, substanțele care au trecut prin aparatul Golgi sunt livrate la adresa, în funcție de etichetele pe care le-au primit în aparatul Golgi.
14. Eps granulat și neted. Structura și caracteristicile funcționării în celule de același tip.
Reticulul endoplasmatic (EPS) - un sistem de canale tubulare comunicante sau separate și cisterne aplatizate situate în întreaga citoplasmă a celulei. Sunt delimitate de membrane (organite membranare). Uneori, rezervoarele au expansiuni sub formă de bule. Canalele EPS se pot conecta cu membrane de suprafață sau nucleare, contact cu complexul Golgi.
În acest sistem se pot distinge EPS neted și aspru (granular).
EPS dur. Pe canalele ER rugoase, ribozomii sunt localizați sub formă de polizomi. Aici are loc sinteza proteinelor, produse în principal de celulă pentru export (scoaterea din celulă), de exemplu, secreții de celule glandulare. Aici are loc formarea lipidelor și proteinelor membranei citoplasmatice și asamblarea acestora. Cisternele strâns împachetate și canalele ER granulare formează o structură stratificată în care sinteza proteinelor este cea mai activă. Acest loc se numește ergastoplasmă.
EPS neted. Nu există ribozomi pe membranele netede ale RE. Aici are loc în principal sinteza grăsimilor și substanțelor similare (de exemplu, hormoni steroizi), precum și a carbohidraților. Prin canalele EPS netede, materialul finit se deplasează, de asemenea, la locul de ambalare în granule (în zona complexului Golgi). În celulele hepatice, ER neted participă la distrugerea și neutralizarea unui număr de substanțe toxice și medicinale (de exemplu, barbiturice). În mușchii striați, tubii și cisternele ER netede depun ioni de calciu.
15. Complexul Golgi. Structură și funcții.
Complexul Golgi este o structură membranară inerentă oricărei celule eucariote. Complexul Golgi este format din cisterne aplatizate, de obicei dispuse în stive (dictiozomi). Rezervoarele nu sunt izolate, ci sunt interconectate printr-un sistem de tuburi. Prima cisternă din nucleu se numește cis-pol al complexului Golgi, iar ultima, respectiv, trans-pol. Numărul de cisterne din diferite celule ale diferitelor organisme poate varia, dar, în general, structura complexului Golgi în toate eucariotele este aproximativ aceeași. În celulele secretoare, este deosebit de puternic dezvoltat. Funcțiile complexului Golgi sunt de a transporta proteinele la destinație, precum și de glicozilare, deglicozilare și modificarea lanțurilor de oligozaharide.
Complexul Golgi este caracterizat de anizotropie funcțională. Proteinele nou sintetizate sunt transportate din reticulul endoplasmatic la polul cis al dictiozomilor folosind vezicule. Apoi se deplasează treptat spre trans-pol, suferind modificări treptate (pe măsură ce se îndepărtează de nucleu, compoziția sistemelor enzimatice din rezervoare se modifică). În cele din urmă, proteinele călătoresc la destinația lor finală în vezicule care înmugurează din polul trans. Complexul Golgi asigură transportul proteinelor în trei compartimente: către lizozomi (precum și către vacuola centrală a celulei vegetale și vacuolele contractile ale protozoarelor), către membrana celulară și către spațiul intercelular. Direcția transferului de proteine este determinată de etichete glicozidice speciale. De exemplu, un marker pentru enzimele lizozomale este manoza-6-fosfat. Maturarea și transportul proteinelor mitocondriale, nucleare și cloroplastice au loc fără participarea complexului Golgi: ele sunt sintetizate de ribozomi liberi și apoi intră direct în citosol. O funcție importantă a complexului Golgi este sinteza și modificarea componentei carbohidrate a glicoproteinelor, proteoglicanilor și glicolipidelor. De asemenea, sintetizează multe polizaharide, cum ar fi hemiceluloza și pectina din plante. Cisternele complexului Golgi conțin o gamă întreagă de glicoziltransferaze și glicozidaze diferite. Ele suferă, de asemenea, sulfatarea reziduurilor de carbohidrați.
Organele - structuri prezente constant în citoplasmă, specializate în îndeplinirea anumitor funcții în celulă. Ele sunt împărțite în organite cu semnificație generală și specială.
Reticulul endoplasmatic
Reticulul endoplasmatic, sau reticulul endoplasmatic, este un sistem de cisterne cu membrană plate și tubuli membranari. Rezervoarele și tubulii cu membrană sunt interconectate și formează o structură membranară cu un conținut comun. Acest lucru vă permite să izolați anumite zone ale citoplasmei de nialoplasma principală și să implementați unele funcții celulare specifice în ele. Rezultatul este diferențierea funcțională. zone diferite citoplasma. Structura membranelor EPS corespunde modelului fluid-mozaic. Din punct de vedere morfologic, există 2 tipuri de EPS: neted (granular) și aspru (granular). ER neted este reprezentat de un sistem de tubuli membranari. Rough EPS este un sistem de rezervoare cu membrană. Pe partea exterioară a membranelor EPS aspre sunt ribozomi. Ambele tipuri de EPS sunt dependente structural - membranele unui tip de EPS pot trece în membrane de alt tip.
Funcțiile reticulului endoplasmatic:
1. ER granular este implicat în sinteza proteinelor, în canale se formează molecule de proteine complexe.
2. Smooth ER este implicat în sinteza lipidelor și carbohidraților.
3. Transport materie organicăîn celulă (prin canalele ER).
4. Împarte celula în secțiuni - în care diferite reacții chimiceși procesele fiziologice.
EPS neted este multifuncțional. În membrana sa există proteine-0 enzime care catalizează reacțiile de sinteză a lipidelor membranare. În ER netedă, sunt sintetizate și unele lipide non-membranare (hormoni steroizi). Compoziția membranei acestui tip de EPS include purtători de Ca 2+. Ei transportă calciul de-a lungul unui gradient de concentrație (transport pasiv). În transportul pasiv, ATP este sintetizat. Cu ajutorul lor, concentrația de Ca 2+ din hialoplasmă este reglată în EPS neted. Acest parametru este important pentru reglarea microtubulilor și microfibrilelor. În celulele musculare, ER neted reglează contracția musculară. În EPS are loc detoxifierea multor substanțe dăunătoare celulei (droguri). ER neted poate forma vezicule membranoase sau microcorpi. Astfel de vezicule desfășoară reacții oxidative specifice izolat de EPS.
functie principala eps dur este sinteza proteinelor. Acest lucru este determinat de prezența ribozomilor pe membrane. Membrana ER rugoasă conține proteine speciale riboforine. Ribozomii interacționează cu riboforinele și sunt fixați pe membrană într-o anumită orientare. Toate proteinele sintetizate în ER au un fragment de semnal terminal. Pe ribozomii ER brut, sunt sintetizate trei tipuri de proteine:
1. Proteinele membranare. Toate proteinele plasmolemei, membranele ER în sine și majoritatea proteinelor altor organite sunt produse ale ribozomilor ER.
2. proteine secretoare . Aceste proteine intră în reticulul endoplasmatic și apoi sunt expulzate din celulă prin exocitoză.
3. Proteine intraorganoide. Aceste proteine sunt localizate și funcționează în cavitățile organelelor membranare: ER însuși, complexul Golgi, lizozomi și mitocondrii. EPS este implicat în formarea biomembranelor.
În cisternele de ER brut, apare modificarea post-translațională a proteinelor.
ER este un organel versatil Celulele eucariote. Încălcarea structurii și funcției EPS duce la consecințe grave. EPS este locul de formare a veziculelor membranare cu funcții specializate ( peroxizomii).
Complexul Golgi.
Complexul Golgi este un organel membranar universal din celulele eucariote. Partea structurală a complexului Golgi este reprezentată de sistem rezervoare cu membrană, formând un teanc de rezervoare. Această stivă se numește dictiozom. Tubulii și veziculele membranare se îndepărtează de ele.
Complexul Golgi poate fi reprezentat într-o celulă printr-un dictiozom într-o secțiune specială a citoplasmei. O celulă poate conține mai mulți dictiozomi izolați interconectați.
În dictiozomul complexului Golgi se disting 2 poli: proximal (regenerativ) și distal (funcțional). Polul proximal este orientat spre citoplasmă sau nucleu, iar polul distal este orientat spre plasmolemă.
Structura membranelor complexului Golgi corespunde structurii fluido-mozaice. Membranele diferiților poli sunt separate prin cantitatea de glicolipide și glicoproteine. La polul proximal se formează noi cisterne de dictiozomi. Veziculele membranoase mici se desprind din EPS neted și se deplasează în zona polului proximal. Aici se unesc și formează o cisterna mai mare. Ca urmare a acestui proces, substanțele care sunt sintetizate în EPS pot fi transportate în rezervoarele complexului Golgi. Veziculele se desprind de pe suprafețele laterale ale polului distal, care sunt implicate în engiocitoză.
Complexul Golgi îndeplinește 3 funcții celulare generale:
Cumulativ
Secretar
Agregare
În cisternele complexului Golgi au loc anumite procese biochimice. Ca urmare, se efectuează modificarea chimică a componentelor membranei rezervoarelor complexului Golgi și a moleculelor din interiorul acestor rezervoare. În membranele cisternelor polului proximal există enzime care realizează sinteza carbohidraților (polizaharide) și atașarea acestora la lipide și proteine, adică. are loc glicozilarea. Prezența acestei sau a altei componente carbohidrate în proteinele glicozilate determină soarta acestora. În funcție de aceasta, proteinele intră în diferite regiuni ale celulei și sunt secretate. Glicozilarea este una dintre etapele de maturare a secretului. În plus, proteinele din cisternele complexului Golgi pot fi fosforilate și acetilate. Polizaharidele libere pot fi sintetizate în complexul Golgi. Unele dintre ele suferă sulfatare cu formarea de mucopolizaharide (glicozaminoglicani). O altă opțiune pentru maturarea secreției este condensarea proteinelor. Acest proces constă în îndepărtarea moleculelor de apă din granulele secretoare, ceea ce duce la compactarea secretului.
De asemenea, universalitatea complexului Golgi în celulele eucariote este participarea acestuia la formare lizozomi.
Lizozomi.
Lizozomi sunt organite membranoase ale celulei. În interiorul lizozomilor se află o matrice lizozomală de mucopolizaharide și enzime proteice.
Membrana lizozomală este un derivat al membranei EPS, dar are propriile sale caracteristici. Aceasta se referă la structura stratului bilipid. În membrana lizozomului, aceasta nu este continuă (nu continuă), dar include micele lipidice. Aceste micelii alcătuiesc până la 25% din suprafața membranei lizozomale. Această structură se numește placă-micelară. O varietate de proteine sunt localizate în membrana lizozomului. Acestea includ enzime: hidrolaze, fosfolipaze; și proteine cu greutate moleculară mică. Hidrolazele sunt enzime specifice lizozomului. Ele catalizează reacțiile de hidroliză (clivaj) substanțelor macromoleculare.
Funcțiile lizozomilor:
1. Digestia particulelor în timpul fagocitozei și pinocitozei.
2. Protectiv în timpul fagocitozei
3. Autofagie
4. Autoliza în ontogenie.
Funcția principală a lizozomilor este participarea la ciclurile heterofagotice (heterofagie) și la ciclurile autofagotice (autofagie). În heterofagie, substanțele străine celulei sunt descompuse. Autofagia este asociată cu descompunerea substanțelor proprii celulei. Varianta obișnuită a heterofagiei începe cu endocitoză și formarea unei vezicule endocitare. În acest caz, vezicula se numește heterofagozom. Pe ER brut, proteinele lizozomului, inclusiv hidrolazele, sunt sintetizate. Ele sunt în primul rând glicozilate ca parte a veziculei membranoase și direcționate către complexul Golgi. Aici are loc glicozilarea suplimentară și formarea unei matrice lizozomale. Ca rezultat, se formează lizozomi primari cu hidrolaze inactive. Inactivarea hidrolazelor se datorează glicozilării lor și acțiunii inhibitorilor specifici de hidrolază. Acești inhibitori sunt proteine membranoase de lizozom cu greutate moleculară mică.
Lizozomul primar converge în heterofagozom cu ajutorul microtubulilor. După contactul acestor vezicule membranoase, ele se apropie una de cealaltă și formează un lizozom secundar sau heterofagolizozom. Formarea unui heterofagozom activează pompa de protoni, care transportă protoni H + în ea. Mediul lizozomului secundar devine mai acid, ceea ce induce activitatea hidrolazelor. Ca urmare, substanța care a intrat în lizozomul secundar este supusă scindării. Produșii de hidroliză cu greutate moleculară mică trec prin membrana lizozomului secundar în hialoplasmă. După ce hidroliza este finalizată, lizozomul secundar poate fuziona cu un nou heterofagozom și poate începe un nou ciclu heterofagotic. După mai multe astfel de cicluri, hidrolazele își pierd activitatea și se transformă într-un telolizozom sau corp rezidual. Telolizomul conține resturi de substanțe nedigerate. Poate fi stocat în hialoplasmă sau inclus în procesul de exocitoză.
Într-o altă variantă de heterofagie, nu există o etapă de endocitoză a substanțelor străine. În acest caz, lizozomul primar este imediat inclus în exocitoză. Ca urmare, hidrolazele matricei se găsesc în glicocalixul celulei și sunt capabile să scindeze substanțele străine extracelulare.
Autofagie simplă (macroautofagie) nu are diferențe fundamentale față de heterofagie. În acest caz, substanța divizată este înconjurată de o secțiune nu a plasmolemei, ci a endomembranei (EPS, complexul Golgi). Ca rezultat, se formează un autofagozom. Se fuzionează cu lizozomul primar pentru a forma lizozomul secundar. Se numește autofagolizozom. Soarta ulterioară a autofagolizozomului este similară cu soarta lizozomului secundar în ciclul heterofagotic. Autofagia distruge organele membranei celulare (mitocondriile). În acest caz, membranele lizozomului primar se îmbină cu membrana exterioară a mitocondriilor. În același mod, conținutul diferitelor vezicule membranare ale citoplasmei poate fi distrus.
O altă variantă a autofagiei a fost numită lizofagie.În acest caz, nu se formează autofagozom. Un caz particular de lizofagie este microautofagia, când o substanță degradabilă trece prin membrana lizozomului în matricea lizozomală. Există o opțiune când autofagia este realizată prin divizarea substanțelor din hialoplasmă în sine. În acest caz, fosforilarea proteinelor membranare primare a lizozomului induce eliberarea de nidrolaze din lizozom în hialoplasmă. Aici are loc hidroliza extra-lizozomală a substanțelor. Multe funcții celulare se bazează pe autofagie și heterofagie.
Autofagia este caracteristică tuturor celulelor eucariote. Când substanțele sunt distruse în procesul de autofagie, se formează metaboliți cu greutate moleculară mică. Sunt folosite în plastic și schimb de energieși îndeplinesc o funcție trofică. Pe baza autofagiei și heterofagiei, lizozomii sunt implicați în procesarea anumitor proteine. Proteina tiroglobulinei leagă iodul și pătrunde în celulele epiteliale ale glandei tiroide prin endocitoză. Aici, lizozomul descompune tiroglobulina în hormoni cu greutate moleculară mică: tiroxina și triiodotironina. În absența activității hidrolazelor lizozomale, catabolismul normal (diviziunea) substanțelor este perturbat. În funcție de încălcarea catabolismului, bolile de acumulare sunt clasificate în: glicogenoze, mucopolizaharidoze, mucolipidoze, sfingolipidoze. Acestea sunt boli ereditare.
Peroxizomii.
Peroxizomii(microcorpii) sunt similare structural cu lizozomii. Ele constau dintr-un matirix și o nucleotidă. Matricea peroxizomilor conține până la 15 enzime. Cele mai importante dintre acestea sunt peroxidaza și catalaza, D-aminoacid oxidaza și uratrooxidaza. Nucleotida peroxizomului corespunde regiunii de condensare a enzimei. Peroxizomii se formează în ER, înmuguriți din ER agranular, enzimele lor sunt parțial sintetizate în ER granular și parțial în hialoplasmă. Membrana peroxizomală este impermeabilă la ioni și la substraturi cu greutate moleculară mică.
Peroxizomii este centrul principal al producerii de oxigen în celulă. Ca urmare a oxidării aminoacizilor, carbohidraților, se formează H 2 O 2 care, datorită catalazei, se descompune în apă și O 2. Peroxizomii mari ai ficatului și rinichilor joacă un rol important în neutralizarea unui număr de substanțe. În plus, sunt implicați în catabolism (în schimbul de aminoacizi, oxalat și poliamine).
În prezent, a fost descoperită o clasă de boli ereditare - boli peroxizomale, a căror dezvoltare se datorează unui defect al peroxizomilor. În aceste boli, organele sunt afectate, se dezvoltă tulburări sistem nervos provocând moartea la pacienții din copilărie.
Mitocondriile.
mitocondriile sunt organite membranare universale ale celulelor. Mitocondriile au 2 membrane - exterioara si interioara. Între aceste membrane există un spațiu intermembranar. În unele părți ale membranei formează locuri de contact. Mitocondriile conțin matricea mitocondrială. Localizează molecule de ADN mitocondrial, ribozomi proprii, ARN, proteine, metaboliți cu greutate moleculară mică.
Membrana exterioară conține mai mult de 80% lipide și mai puțin de 20% proteine, în timp ce membrana interioară conține opusul. Printre proteinele membranei exterioare sunt porine care formează pori. Prin intermediul lor, din hialoplasmă provin molecule de o anumită dimensiune. Ca urmare, membrana exterioară are o permeabilitate nespecifică. Receptorii speciali și proteinele canalului sunt localizați în zona locurilor de contact. Membrana interioară formează crestae. Pe ele, din partea matricei mitocondriale, sunt localizate corpuri în formă de ciupercă - componente proteice care realizează sinteza ATP.
Simptomele majorității bolilor mitocondriale apar odată cu vârsta, ceea ce se datorează probabil acumulării de mutații efectuate de H 2 O 2 și O 2 . pentru că aceste substante sunt generate in cantitati maxime in timpul fosforilarii oxidative, organele care au cel mai mult nevoie de energie mitocondriala (SNC, inima, muschii scheletici, rinichi, ficat, insulițe Langerhans).
Ciclul de viață al mitocondriilor este de aproximativ 10 zile, distrugerea lor are loc prin autofagie, iar organele muritoare sunt înlocuite cu altele noi, care se formează prin relansarea celor anterioare. Replicarea ADN-ului mitocondrial are loc în orice fază a ciclului celular, indiferent de ADN-ul nuclear.
Funcții mitocondriale:
1. Centrul respirator și energetic al celulei - absorb oxigenul necesar pentru a treia etapă (aerobă) de disimilare.
2. Sinteza lor ADN, ARN, părți de proteine.
Ribozomi.
Ribozomi– organele sens general care nu au o structură membranară. locul sintezei proteinelor. D=15-35 nm. Se găsesc în citoplasmă, plastide și mitocondrii. Majoritatea ribozomilor se formează în nucleolul nucleului - sub formă de 2 subunități care părăsesc nucleul și se combină pentru a forma un ribozom, care constă dintr-o subunitate mare și mică. Fiecare subunitate conține ARNr și o proteină.
Ribozomii, care se conectează cu ARNm în timpul sintezei proteinelor, formează polizomi (poliribozomi).
Ribozomii sunt asociați cu ER granular, sintetizează proteine secretoare în mod normal sau rămân în membranele celulei.
Funcțiile ribozomului - sinteza proteinei.
Plastide.
plastide - Organele de importanță generală în celulele vegetale, Euglena verde (protozoare). Se disting: cloroplaste, cromoplaste, leucoplaste.
Cloroplaste - plastide verzi înconjurate de două membrane. Strat interior membranele din cavitatea cloroplastului formează saci plate - tilocoizi. Au formă de disc, formează un teanc de ≈ 50 de bucăți, se numesc stivele granule. Cloroplastul conține 40-60 de granule. Spațiul dintre tilacoizi este umplut cu stroma (matricea) cloroplastei din proteine, lipide, carbohidrați, enzime, ATP, ADN, ARN, ribozomi. Cloroplastele se formează din proplastide - corpuri mici nediferențiate. Cloroplastele se reproduc prin diviziune. Cloroplastele se pot transforma in cromoplaste si leucoplaste toamna.
Funcțiile cloroplastelor:
1. Fotosinteza
2. Sinteza proteinelor proprii.
leucoplaste - plastide incolore în părțile necolorate ale plantelor: celule, endosperm de semințe, tuberculi, rădăcinoase. Acestea sunt organite cu două membrane, în interiorul a 2-3 excrescențe. Forma este rotunjită. Ele trec în cloroplaste și cromoplaste.
Funcţie:
1. Acumularea de nutrienți - amidon, grăsimi, proteine.
Cromoplaste - plastide cu două membrane de formă filamentoasă, lamelară sau altă formă. Culoare galben-roșu-maro-portocaliu datorită pigmenților carotenoide. Se găsește în celulele fructifere. Cromoplastele sunt etapa finală în dezvoltarea plastidelor - cloroplastele și leucoplastele se transformă în ele.
Funcţie:
1. În celulă: joacă rolul unui fel de filtru de lumină pentru cloroplaste în procesul de fotosinteză; loc de sinteza si localizare a pigmentilor vegetali.
2. Culoarea corolelor florilor - atragerea insectelor polenizatoare.
3. Colorarea fructelor - atracția animalelor - dispersarea semințelor.
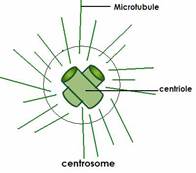
Centrul celular.
Centrul celular(centrizom) - un organoid al unei structuri non-membranare în celulele animalelor și plantelor inferioare. Este situat în apropierea nucleului, este format din 2 centrioli - corpuri cilindrice lungi de 500 nm, situate perpendicular unul pe celălalt. Pereții sunt formați din 9 tripleți de microtubuli. Centrizomul este înconjurat de o citoplasmă mai ușoară, centrosferă.
Funcţie:
Centru de formare a microtubulilor fusului. În timpul diviziunii celulare, centrizomul este împărțit în 2 părți și un centrisol se deplasează la un pol al celulei, celălalt la celălalt și formează fusuri de diviziune, asigurând o distribuție uniformă a cromozomilor între celulele fiice.
EPS este un organel cu o singură membrană care este o colecție de vacuole membranare, tubuli și saci plate (cisternă) distribuite într-un fel sau altul în citoplasmă.
Parte principală:
EPS neted (granular) - aproape nu există cisterne, nicio structură vizibilă nu este asociată cu membranele vacuolelor și tubilor. Funcții: Enzimele de hidroxilare (sau oxidarea microzomală) sunt încorporate în membranele EPS netede. Sunt necesare - în sinteza multor lipide (de exemplu, hormoni steroizi) - și pentru neutralizarea compușilor toxici. EPS neted este deosebit de pronunțat în celulele glandelor suprarenale și gonadelor, care sintetizează hormoni steroizi, și ficatului, în legătură cu sinteza colesterolului.
 10. Centrozom. Axul de diviziune. Structura și funcția moleculară.
10. Centrozom. Axul de diviziune. Structura și funcția moleculară.
Scurtă recenzie:
Centrul celular (centrozom)- un organel non-membranar, principalul centru de organizare a microtubulilor și un regulator al ciclului celular în celulele eucariote.
Axul de diviziune
Raspuns complet:
Centrioli, de obicei dispuși într-o pereche (diplozom) și înconjurați de o zonă de citoplasmă mai ușoară, din care se extind radial fibrile subțiri (centrosferă). Combinația de centrioli și centrosferă se numește centru celular. Combinația de centrioli și centrozom se numește centru celular.
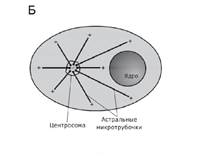 Centrul celular (centrozom)- un organel non-membranar, principalul centru de organizare a microtubulilor și un regulator al ciclului celular în celulele eucariote. Rolul său important este în diviziunea celulară, în special, în formarea fusului mitotic. Descoperit pentru prima dată în 1883 de Theodore Boveri, care l-a numit „un organ special diviziune celulara. Celula conține unul sau doi centrozomi.
Centrul celular (centrozom)- un organel non-membranar, principalul centru de organizare a microtubulilor și un regulator al ciclului celular în celulele eucariote. Rolul său important este în diviziunea celulară, în special, în formarea fusului mitotic. Descoperit pentru prima dată în 1883 de Theodore Boveri, care l-a numit „un organ special diviziune celulara. Celula conține unul sau doi centrozomi.
 Fiecare centriol este construit din 27 de elemente cilindrice (microtubuli) grupate în 9 triplete formate ca urmare a polimerizării proteinei tubulinei. Nouă triplete de microtubuli sunt aranjate într-un cerc, formând astfel un cilindru gol.
Fiecare centriol este construit din 27 de elemente cilindrice (microtubuli) grupate în 9 triplete formate ca urmare a polimerizării proteinei tubulinei. Nouă triplete de microtubuli sunt aranjate într-un cerc, formând astfel un cilindru gol.
În interfaza ciclului celular, centrozomii sunt asociați cu membrana nucleară. În profaza mitozei, membrana nucleară este distrusă, centrozomul se divide, iar produșii diviziunii sale (centrozomii fiice) migrează către polii nucleului de divizare. Microtubulii care cresc din centrozomi fiice sunt atașați la celălalt capăt de așa-numitele kinetocore de pe centromerii cromozomilor, formând un fus de diviziune. La sfârșitul diviziunii, fiecare dintre celulele fiice conține un singur centrozom.
Pe lângă participarea la diviziunea nucleară, centrozomul joacă un rol important în formarea flagelilor și a cililor. Centriolii localizați în acesta acționează ca centre de organizare pentru microtubulii axonemelor flagelului. La organismele lipsite de centrioli (de exemplu, marsupiale), flagelii nu se dezvoltă.
Axul de diviziune- o structură dinamică care se formează în mitoză și meioză pentru a asigura segregarea cromozomilor și diviziunea celulară. 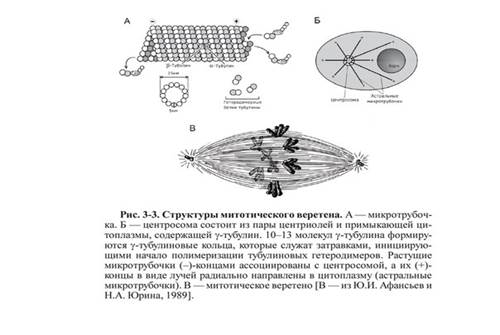 Un fus tipic este bipolar - între cei doi poli se formează un sistem de microtubuli în formă de fus. Microtubulii fusului se atașează de cinetocorele cromatide la centromeri și asigură mișcarea cromozomilor spre poli.
Un fus tipic este bipolar - între cei doi poli se formează un sistem de microtubuli în formă de fus. Microtubulii fusului se atașează de cinetocorele cromatide la centromeri și asigură mișcarea cromozomilor spre poli.
Fusul este format din trei elemente structurale principale: microtubuli, poli de diviziune și cromozomi. La animale, centrozomii care conțin centrioli sunt implicați în organizarea polilor de diviziune. Un rol important în formarea fusului îl au proteinele motorii aparținând familiilor dineinelor și kinezinelor.
Un fus de fisiune complet se formează în stadiul de prometafază după distrugerea membranei nucleare.
Fusul de diviziune al unei celule tipice de mamifer este format din trei elemente structurale- centrozomi, microtubuli și cromozomi - care formează o structură bipolară simetrică. La polii fusului se află centrozomi, organele mici care funcționează ca centre de organizare a microtubulilor. Fiecare centrozom constă dintr-o pereche de centrioli înconjurați de multe proteine diferite. Între polii fusului se află cromozomi condensați, formați dintr-o pereche de cromatide, fixați la centromer. Pe regiunile centromerice ale cromozomilor se află cinetocorii - structuri complexe responsabile de atașarea la microtubulii fusului.
Axul de divizare este format din două semifusuri. Semifusul este format din microtubuli polarizați. Capetele minus negative ale microtubulilor se adună la polii fusului în jurul centrozomilor. Capetele plus ale microtubulilor se îndepărtează de cei doi poli și se intersectează în partea ecuatorială mijlocie a fusului. La majoritatea vertebratelor, semifusul este format din 600-750 de microtubuli, dintre care 30-40% se termină în cinetocori. Se numesc microtubulii care leagă polii fusului de cinetocorii cromozomilor kinetocor. Mai mult, fiecare kinetocor, în timpul formării fusului, este asociat cu mulți microtubuli și formează un mănunchi cinetocor. Se numesc microtubuli care se află între poli și nu se atașează de kinetocori interpolară. O parte din microtubulii fusului formează structuri radiale în jurul fiecărui pol, numite stele sau asteri. Acești microtubuli se numesc astral.
Întrebarea #11
mitocondriile
Scurtă trecere în revistă (din manual) - Formațiuni cu membrană dublă care au propriul ADN se presupune că au apărut din procariote după combinarea cu celulele eucariote ca urmare a evoluției și coexistenței ulterioare cu acestea (simbioză). Ele asigură sinteza ATP datorită reacțiilor de fosforilare oxidativă. Mitocondriile controlează conținutul intracelular al ionilor de calciu, asigură generarea de căldură, precum și moartea celulară programată.
Partea principală (de pe Internet) - Acestea sunt organele de mărimea unei bacterii. Mitocondriile sunt limitate la două membranelor - exterior netedȘi pliat pe interior, care are o suprafață foarte mare. Pliurile membranei interioare intră adânc în matricea mitocondrială, formând un sept transversal - cristae. Spațiul dintre membranele exterioare și interioare este denumit în mod obișnuit ca spaţiul intermembranar. Membranele mitocondriale conțin proteine membranare integrale. Membrana exterioară conține porine, care formează pori și fac membranele permeabile la substanțe cu o greutate moleculară de până la 10 kDa. Membrana interioară a mitocondriilor este impermeabilă la majoritatea moleculelor; excepția este O 2, CO 2, H 2 0. Membrana interioară a mitocondriilor este caracterizată printr-un conținut neobișnuit de mare de proteine (75%). proteine de transport, enzime, componente ale lanțului respiratorȘi ATP sintetaza. În plus, conține un fosfolipid neobișnuit cardiolipină. Matricea este, de asemenea, îmbogățită în proteine, în special în enzime ale ciclului citratului.
funcții metabolice Mitocondriile sunt "centrala electrica" celulele, deoarece datorită degradării oxidative a nutrienților, acestea sintetizează cea mai mare parte din ATP (ATP) necesar celulei. Următoarele procese metabolice sunt localizate în mitocondrii: conversia piruvatului în acetil-CoA catalizată de complexul piruvat dehidrogenază: ciclu de citrat; lanțul respirator, asociat cu sinteza ATP(combinația acestor procese se numește „ fosforilarea oxidativă»); descompunerea acizilor grași prin β-oxidareși parțial ciclul ureei. De asemenea, mitocondriile furnizează celulei cu produse ale metabolismului intermediar și acționează, împreună cu ER, ca depozit de ioni de calciu, care, cu ajutorul pompelor ionice, menține concentrația de Ca 2+ din citoplasmă la un nivel scăzut constant (sub 1 µmol/l). Funcția principală a mitocondriilor este captarea substraturilor bogate în energie(acizi grași, piruvat, scheletul carbonic al aminoacizilor) din citoplasmă și clivajul lor oxidativ cu formarea de CO 2 și H 2 O, cuplat cu sinteza ATP.
12. Ribozomi. Poliribozomi. Ribozomi mitocondriali.
Scurtă recenzie:
Un ribozom este un organel non-membranar cu două subdiviziuni (subunitate mică și mare) format din ARNr și proteine. Ribozomii au formă sferică sau ușor elipsoidală.
Parte principală:
Un ribozom este un organel non-membranar cu două subdiviziuni (subunitate mică și mare) format din ARNr și proteine. Ribozomii au formă sferică sau ușor elipsoidală. Servește la biosinteza proteinelor din aminoacizi . Ribozomii sunt împărțiți în ribozomi liberi și cei asociați cu membranele ER și membrana nucleară exterioară. Ribozomii liberi sintetizează proteine pentru celula însăși și leagă ribozomii pentru export. Sinteza ribozomilor la eucariote are loc într-o structură intranucleară specială - nucleolul.
Un polizom, sau poliribozom, este mai mulți ribozomi care traduc simultan o moleculă de ARNm.
Ribozomii mitocondriali sau mitoribozomii sunt asociați cu matricea mitocondrială.
Întrebarea #13
aparate Golgi
Scurtă recenzie.
Aparatul Golgi este un sistem de structuri membranare intracelulare: rezervoare și vezicule în care se acumulează substanțe sintetizate pe membranele EPS.
Substanțele sunt livrate în complexul Golgi în vezicule membranoase care se desprind de reticulul endoplasmatic și se unesc cu cisternele complexului Golgi. Aici, aceste substanțe suferă diferite transformări biochimice și apoi sunt din nou împachetate în vezicule membranare, iar cele mai multe dintre ele sunt transportate în membrana citoplasmatică. Membrana veziculelor fuzionează cu membrana citoplasmatică, iar conținutul este expulzat în afara celulei prin exocitoză.
În complexul Golgi de celule vegetale, sunt sintetizate polizaharidele peretelui celular (cochilie).
O altă funcție importantă a complexului Golgi este formarea lizozomilor.
Complexul Golgi a fost descoperit în 1898 de histologul italian Camillo Golgi.În celulele nervoase.
Parte principală.
Complexul Golgi este un loc de condensare și acumulare a produșilor de secreție produși în alte părți ale celulei, în principal în EPS.
Pe partea exterioară, concavă, a stivei de bule, se formează în mod constant cisterne noi, iar pe interiorul rezervoarelor se transformă înapoi în bule.
Sub microscopie cu lumină, poate fi distribuit sub formă de rețele complexe sau zone separate situate difuz (dictiozomi). Forma și poziția organelului nu au o importanță fundamentală și se pot schimba în funcție de starea funcțională a celulei.
Microscopia electronică arată că complexul este format din grupuri de cisterne plate
În general, complexul Golgi este implicat în segregare - aceasta este separarea, separarea anumitor părți de masa principală și acumularea de produse sintetizate în EPS, în rearanjamentele lor chimice, maturare.
Funcția secretorie a complexului Golgi este că proteina exportată sintetizată pe ribozomi, care este separată și acumulată în interiorul rezervoarelor EPS, este transportată în vacuolele aparatului lamelar.
Complexul Golgi poate crește dramatic în dimensiune în celulele care realizează activ funcția secretorie, de obicei însoțită de dezvoltarea EPS, iar în cazul sintezei proteinelor, nucleolul.
În timpul diviziunii celulare, complexul Golgi se descompune în cisterne sau vezicule individuale, care sunt distribuite între două celule în diviziune și, la sfârșitul telofazei, restabilesc integritatea structurală a organelului.
În afara diviziunii, există o reînnoire continuă a aparatului membranar datorită veziculelor care migrează din EPS și cisterne distale ale dictiozomului datorită compartimentelor proximale.
despre conținut în timpul exocitozei lizozomale.
14. lizozomi- structurile unimembranare se formează prin fuziunea endozomilor perinucleari care conțin hidrolaze lizozomale și proteine membranare lizozomale cu vezicule de degradat (endozom periferic, fagozom sau vacuola autofagocitară).
* endozomi perinucleari sunt formate prin fuziunea veziculelor care conțin hidrolaze lizozomale după sinteza lor în eps granular și prelucrare în complexul Golgi, și vezicule, în membrana cărora sunt construite proteine specifice membranei lizozomale.
* endozomii periferici format prin endocitoză.
*corpuri multiveziculare format prin fuziunea endozomilor perinucleari si periferici.
*fagolizozom format prin fuziunea endozomului perinuclear cu fagozomul.
*autofagolizozom se formează prin fuziunea endozomului perinuclear și a unei vacuole autofagocitare care conține molecule endogene și organele de degradat.
* corpuri reziduale- lizozomi de orice tip care contin material nedigerat (lipofuscina, hemosiderina).
Funcţie:
Cataliza hidrolitică (în mediu acvatic) despicare acizi nucleici, proteine, grăsimi, polizaharide și mucopolizaharide, altele compuși chimici la valori scăzute ale pH-ului. În termeni citobiologici - digestia intracelulară a substanțelor și structurilor.
Peroxizomi - organite cu o singură membrană, vezicule cu miez dens în electroni. Membrana organelelor conține proteine specifice - peroxine, iar matricea conține peste 40 de enzime care catalizează procesele anabolice (biosinteza acizilor biliari, H2O3, respirație dependentă, degradarea xenobioticelor).
Reticul endoplasmatic (reticul endoplasmatic)- un organel al unei celule eucariote. Sub forma unei rețele de canale și rezervoare, limitate de o singură membrană, se ramifică pe întregul volum al citoplasmei. Reticulul endoplasmatic este implicat în metabolism: sintetizează lipide pentru membrana dublă exterioară a celulei și pentru membrana proprie, unică, asigură transportul de substanțe între organele celulare, servește drept tezaur de substanțe și loc pentru izolarea acestora.
Există două tipuri de reticul endoplasmatic - aspru și neted. Rețeaua aspră poartă numeroși ribozomi pe suprafața exterioară. Proteinele sintetizate pe ele sunt izolate aici de alte proteine ale celulei prin transferul lor prin membrana canalului reticulului endoplasmatic. Ea „recunoaște proteinele trecute după” semnalul lor special „termină. Scindarea acestor capete după trecerea proteinei prin membrană se numește maturare proteică. Unele proteine – secretoare – sunt eliberate din celulă. Altele sunt incluse în toate celulele. membranelor.
Reprezentare schematică nucleul celular, reticulul endoplasmatic și complexul Golgi.
(1) Nucleul celular. (2) Porii membranei nucleare. (3) Reticulul endoplasmatic granular.
(4) Reticulul endoplasmatic agranular. (5) Ribozomi de pe suprafața granularului
reticulul endoplasmatic. (6) Macromolecule (7) Vezicule de transport.
(8) Complexul Golgi. (9) Cis-Golgi (10) Trans-Golgi (11) Cisterne Golgi
O rețea netedă constă din tuburi, canale și bule cu o secțiune transversală mai mică decât într-o rețea brută. Funcțiile sale sunt, de asemenea, diverse: aici se sintetizează lipidele membranare, dar, pe lângă acestea, lipidele nemembranare (de exemplu, hormoni speciali animale), substanțele toxice sunt neutralizate de complexe enzimatice speciale, iar ionii se acumulează. Deci, în mușchii striați, rețeaua netedă servește ca un rezervor de ioni de calciu. Membranele acestei rețele conțin „pompe” puternice de calciu care transportă o cantitate mare de ioni de calciu în orice direcție în sutimi de secundă. În celule specializate rețea lină este diferit, ceea ce este asociat cu funcțiile sale specifice în metabolismul intracelular.
Reticulul endoplasmatic este foarte vulnerabil la impact: pierde rapid ribozomii și se prăbușește. Cu toate acestea, datorită capacității de a reconstrui rapid, se poate recupera.













