Cum este stocată informația genetică în moleculele de ADN? Stocarea și implementarea informațiilor ereditare
1. Potrivire:
1-c, 2-a, 3-b
2. Ce este informația ereditară? Cum este stocată informația genetică în celule?
Informațiile despre structura și viața atât a fiecărei celule individuale, cât și a întregului organism multicelular sunt conținute în secvența nucleotidelor ADN. Aceste informații se numesc ereditare sau genetice. Secțiunea moleculei de ADN care conține informații despre structura primară a unei proteine se numește genă.
3. Caracterizați proprietățile cod genetic
1 - Codul genetic este triplet, adică fiecare aminoacid este codificat de un triplet (codon) - o combinație de trei nucleotide consecutive. 2 - Codul nu se suprapune - aceeași nucleotidă nu poate face parte simultan din două triplete învecinate. 3 - Codul nu este ambiguu - fiecare triplet codifică doar un aminoacid. 4 - În acest sens, codul este redundant (degenerat) - același aminoacid poate fi codificat de mai multe triplete diferite. la un triplet - AUG, triptofan - doar triplet UGG. 5 - Codul este continuu sau, cu alte cuvinte, nu are semne de punctuație. Aceasta înseamnă că, dacă o nucleotidă cade, atunci când o citiți, cea mai apropiată nucleotidă din codonul vecin îi va lua locul. 6 - codul este universal - la toate organismele vii, aceiași aminoacizi corespund acelorași tripleți.
4. Ce sunt codonii de terminare? Care este rolul codonului de start AUG?
Tripleții ARNm (UAA, UAG, UGA) nu codifică aminoacizi. Aceștia sunt așa-numiții codoni de terminare (codoni de oprire). Ele servesc drept semnal pentru sfârșitul sintezei proteinelor. Codonul de pornire vă permite să începeți citirea corectă a codului din ARNm. În molecula de ARNm, codonul de început este AUG, de la acesta începe citirea.
5. De ce un aminoacid este codificat nu de una sau două, ci de trei nucleotide consecutive?
Există 20 de aminoacizi naturali care alcătuiesc proteinele. În același timp, există doar 4 tipuri de reziduuri de nucleotide. Este clar că o nucleotidă nu poate codifica un aminoacid (4 față de 20). Pot exista 16 combinații posibile de două nucleotide (dublete), ceea ce, din nou, nu este suficient pentru a codifica 20 de aminoacizi. Prin urmare, numărul minim de resturi care codifică un aminoacid poate fi trei, adică aminoacizii trebuie să fie codificați de tripleți de nucleotide. Numărul total de tripleți posibili este de 64, ceea ce este mai mult decât suficient pentru a codifica 20 de aminoacizi.
6. O moleculă de ARNm începe cu următoarea secvență de nucleotide: AUGGUATZUUGGZATS... Cu ce secvență de resturi de aminoacizi începe proteina codificată de acest ARNm?
Met-Val-Pro-Trip-Gis
7. Ce crezi semnificație biologică are proprietatea de degenerare a codului genetic?
Degenerarea codului genetic oferă șansa ca, chiar dacă o nucleotidă cade dintr-un triplet, acest triplet poate încă codifica proteina necesară.
8. Acțiunea unor factori ( raze X, radiații ultraviolete, definite substanțe chimice etc.) sau erorile care apar uneori în timpul replicării pot duce la modificarea secvenței nucleotidelor ADN. În acest caz, se poate produce inserția sau pierderea de nucleotide sau înlocuirea unei nucleotide cu alta. În opinia dumneavoastră, ce schimbări duc adesea la perturbarea funcționării normale a celulelor (și chiar la moartea lor) - inserția și pierderea sau înlocuirea nucleotidelor ADN? Justificati raspunsul.
Toate aceste modificări pot duce la o întrerupere a funcționării normale a celulei, deoarece un triplet nou format poate codifica un aminoacid complet diferit.
Informația genetică este un program de proprietăți ale unui organism, primit de la strămoși și încorporat în structuri ereditare sub forma unui cod genetic.
Se presupune că formarea informaţiei genetice a urmat schema: procese geochimice - formare minerală - cataliză evolutivă (autocataliza).
Aproape toată informația genetică este stocată în nucleul celulei. Să ne uităm la ce este și sub ce formă este.
ADN-ul este responsabil de informația genetică, iar în cazul virușilor, ARN-ul. În nucleu, ADN-ul este „pliat” în structuri numite cromozomi. Corpul uman conține peste 2 metri de ADN. Informațiile despre structura proteinelor sunt criptate pe moleculele de ADN și ARN cu un cod genetic special. Această informație este transmisă în procesul de replicare (dublare) ADN-ului. Primim informații genetice la naștere de la mamă și tată sub formă de multe gene. Interesant este că toate celulele din corpul nostru conțin aceeași informație genetică. Atunci cum este posibil să diferite celule funcții complet diferite? Faptul este că toată informația genetică nu este realizată în celule, ci doar în secțiunile necesare - gene.
Este posibil ca primele gene primitive să fi fost cristale microcristaline de argilă, iar fiecare nou strat de argilă să se alinieze în conformitate cu caracteristicile structurale ale celui precedent, ca și cum ar primi informații despre structura de la acesta.
Realizarea informației genetice are loc în procesul de sinteză a moleculelor proteice cu ajutorul a trei ARN: informațional (ARNm), de transport (ARNt) și ribozomal (ARNr). Procesul de transfer al informaţiei trece: - prin canalul de comunicare directă: ADN - ARN - proteină; și - prin canalul de feedback: mediu - proteină - ADN.
Organismele vii sunt capabile să primească, să stocheze și să transmită informații. Mai mult, organismele vii tind să folosească cât mai eficient informațiile primite despre ele însele și despre lumea din jurul lor. Informațiile ereditare încorporate în gene și necesare unui organism viu pentru existență, dezvoltare și reproducere sunt transmise de la fiecare individ către descendenții săi. Aceste informații determină direcția de dezvoltare a organismului, iar în procesul de interacțiune a acestuia cu mediul, reacția la individul său poate fi distorsionată, asigurând astfel evoluția dezvoltării descendenților. În procesul de evoluție a unui organism viu, apar și sunt amintite noi informații, inclusiv valoarea informațiilor pentru aceasta crește.
În timpul implementării informații ereditareîn anumite condiții de mediu, fenotipul organismelor unui dat specii.
Informațiile genetice determină structura morfologică, creșterea, dezvoltarea, metabolismul, depozitul mental, predispoziția la boli și defecte genetice ale organismului.
Mulți oameni de știință, subliniind pe bună dreptate rolul informației în formarea și evoluția viețuitoarelor, au remarcat această împrejurare ca fiind unul dintre criteriile principale ale vieții. Deci, V.I. Karagodin crede: „Viul este o astfel de formă de existență a informațiilor și a structurilor codificate de aceasta, care asigură reproducerea acestor informații în condiții de mediu adecvate”. Legătura informației cu viața este remarcată și de A.A. Lyapunov: „Viața este o stare foarte ordonată a materiei care folosește informațiile codificate de stările moleculelor individuale pentru a dezvolta reacții persistente”. Cunoscutul nostru astrofizician N.S. Kardashev subliniază, de asemenea, componenta informațională a vieții: „Viața apare datorită posibilității de a sintetiza un tip special de molecule care sunt capabile să-și amintească și să folosească la început cele mai simple informații despre mediu inconjuratorși propria lor structură, pe care o folosesc pentru autoconservare, pentru reproducere și, ceea ce este deosebit de important pentru noi, pentru a obține și mai multe informații.” Ecologul F. Tipler atrage atenția asupra acestei capacități a organismelor vii de a stoca și transmite informații. în cartea sa Fizica nemuririi: „Definesc viața ca un fel de informație codificată care este păstrată de selecția naturală.” Mai mult, el crede, dacă acesta este cazul, atunci sistemul de informații despre viață este etern, infinit și nemuritor.
Descoperirea codului genetic și stabilirea legilor biologiei moleculare au arătat nevoia de a combina genetica modernă și teoria darwiniană a evoluției. Astfel, a luat naștere o nouă paradigmă biologică - teoria sintetică a evoluției (STE), care poate fi considerată deja ca biologie neclasică.
Principalele idei ale evoluției lui Darwin cu triada sa - ereditate, variabilitate, selecție naturală - în vedere modernă Evoluția lumii vii este completată de idei nu doar de selecție naturală, ci și de o astfel de selecție, care este determinată genetic. Începutul dezvoltării evoluției sintetice sau generale poate fi considerat opera lui S.S. Chetverikov despre genetica populației, în care s-a demonstrat că nu trăsăturile individuale și indivizii sunt supuși selecției, ci genotipul întregii populații, ci se realizează prin trăsăturile fenotipice ale indivizilor individuali. Acest lucru duce la răspândirea unor schimbări benefice la nivelul populației. Astfel, mecanismul evoluției este implementat atât prin mutații aleatorii la nivel genetic, cât și prin moștenirea celor mai valoroase trăsături (valoarea informației!), care determină adaptarea trăsăturilor mutaționale la mediu, oferind descendenții cei mai viabili. .
Schimbările climatice sezoniere, diversele dezastre naturale sau provocate de om, pe de o parte, duc la o modificare a frecvenței de repetare a genelor în populații și, ca urmare, la o scădere a variabilității ereditare. Acest proces este uneori numit derivă genetică. Și pe de altă parte, la modificări ale concentrației diferitelor mutații și o scădere a diversității genotipurilor conținute în populație, ceea ce poate duce la schimbări în direcția și intensitatea selecției.
Cod genetic este un sistem de înregistrare a informațiilor genetice dintr-o moleculă acid nucleic despre structura moleculei polipeptidice, și anume, numărul, secvența și tipurile de aminoacizi. O genă conține informații despre un lanț polipeptidic, de ex. despre structura primară a proteinei.
Codul genetic se caracterizează prin tripletitate, adică. trei nucleotide în secvență dintr-un lanț de acid nucleic (ADN sau ARN) formează un triplet sau codon (cuvânt de cod) care codifică un aminoacid și locația acestuia în lanțul peptidic. Codonii diferă prin secvența și tipurile de nucleotide (baze azotate). Există 64 de tipuri de codoni, ceea ce corespunde numărului de combinații posibile de 4 (4 tipuri de nucleotide care diferă în baze azotate) cu 3 (43). 61 dintre ei sunt codoni informativi, definesc (codifică) aminoacizi. 3 codoni (în ADN - ATT, ATC, ACT, respectiv, în ARNm - UAA, UAG, UGA) se numesc codoni stop, ei asigură sfârșitul sintezei lanțului proteic. Codonul TAC din ADN sau AUG din ARNm (codifică aminoacidul metionină) este cel de pornire, adică. este primul din gena și sinteza peptidelor începe cu ea.
La descifrarea codului genetic, s-a dovedit că majoritatea aminoacizilor sunt codificați de mai mulți codoni diferiți, cu alte cuvinte, există codoni - sinonime care diferă adesea doar în nucleotide a treia (baze azotate). De exemplu, codonii din ADN CGA, CHG, CHT codifică alanină și codonii pentru HCA, HCH, GCT, GCC, TCT, TCC codifică arginină. Această proprietate a codului genetic se numește degenerare sau redundanță.
Cu toate acestea, s-a demonstrat că un codon codifică doar un aminoacid, adică. poate conține informații despre un singur aminoacid - cu alte cuvinte, codul genetic nu este ambiguu.
Codul genetic nu se suprapune, ceea ce înseamnă că codonii sunt aranjați liniar, iar o nucleotidă face parte dintr-un singur codon; și continuitate - codonii nu sunt separați unul de altul, ei sunt localizați în lanțul de acid nucleic unul după altul, adică. distanța dintre codoni corespunde distanței dintre nucleotide și nu există semnale care să indice începutul sau sfârșitul codonilor.
Universalitatea codului genetic implică faptul că codul genetic al tuturor organismelor se caracterizează prin aceleași proprietăți (tripletitate, degenerare etc.); și că semnificația codonilor este aceeași în toate organismele (cu excepția unor codoni din mitocondrii și bacterii).
În toate organismele procariote și eucariote, informația genetică este înregistrată într-o singură catenă de ADN, care este numită codogenă (informativă sau semnificativă) și este indicată prin semnul „+”, a doua catenă nu poartă informații genetice - non-codogenă ( neinformativ sau nesemnificativ) și este indicat prin semnul „-” .
Păstrarea informațiilor genetice
Cum, atunci, în globulele roșii ale unei persoane sănătoase se formează milioane de molecule identice de hemoglobină, de regulă, fără o singură greșeală în aranjarea aminoacizilor? De ce toate moleculele de hemoglobină din eritrocitele pacienților cu anemie falciformă au aceeași eroare în același loc?
Pentru a răspunde la aceste întrebări, să luăm exemplul tipografiei. De exemplu, a fost publicată o carte N copii. Tot N cărți sunt tipărite dintr-un șablon - o matrice tipografică, deci sunt exact aceleași. Dacă o eroare s-a strecurat în matrice, atunci aceasta ar fi reprodusă în toate cazurile. Rolul matricei în celulele organismelor vii este îndeplinit de moleculele de ADN. ADN-ul fiecărei celule poartă informații nu numai despre proteinele structurale care determină forma celulei (amintiți-vă de eritrocitul), ci și despre toate proteinele enzimatice, proteinele hormonale și alte proteine.
Carbohidrații și lipidele se formează în celulă ca urmare a complexului reacții chimice, fiecare dintre acestea fiind catalizat de propria sa proteină enzimatică. Deținând informații despre enzime, ADN-ul programează structura altor compuși organici, precum și controlează procesele de sinteza și scindarea acestora.
Deoarece moleculele de ADN sunt șabloanele pentru sinteza tuturor proteinelor, ADN-ul conține informații despre structura și activitatea celulelor, despre toate caracteristicile fiecărei celule și ale organismului în ansamblu.
Fiecare proteină este reprezentată de unul sau mai multe lanțuri polimerice. Secțiunea moleculei de ADN care servește ca șablon pentru sinteza unui lanț polipeptidic, adică, în cele mai multe cazuri, o proteină, se numește genă. Fiecare moleculă de ADN conține multe gene diferite. Toate informațiile conținute în moleculele de ADN se numesc genetice. Ideea că informația genetică este înregistrată la nivel molecular și că sinteza proteinelor se desfășoară conform principiului matricei a fost formulată pentru prima dată în anii 1920 de remarcabilul biolog rus N.K. Koltsov.
Descifrarea codului genetic uman
În mai 2006, oamenii de știință care lucrează pentru a descifra genomul uman au publicat o hartă genetică completă a cromozomului 1, care a fost ultimul cromozom uman secvențial incomplet.
O hartă genetică umană preliminară a fost publicată în 2003, marcând finalul oficial al Proiectului genomului uman. În cadrul său, au fost secvențiate fragmente de genom care conțin 99% din genele umane. Precizia identificării genelor a fost de 99,99%. Cu toate acestea, la sfârșitul proiectului, doar patru dintre cei 24 de cromozomi au fost complet secvențiați. Cert este că, pe lângă gene, cromozomii conțin fragmente care nu codifică nicio trăsătură și nu sunt implicate în sinteza proteinelor. Rolul pe care îl joacă aceste fragmente în viața organismului este încă necunoscut, dar tot mai mulți cercetători sunt înclinați să creadă că studiul lor necesită cea mai mare atenție.
Partea finală a lucrării de secvențiere a genomului uman a durat oamenilor de știință aproximativ trei ani. Cromozomul 1 a durat cel mai mult pentru a descifra, deoarece este cel mai lung cromozom din întregul genom. Este de șase ori mai lung decât cei mai scurti cromozomi (21, 22 și Y). Conține aproximativ 8% din codul genetic: 3141 gene și 991 pseudogene, cu multe secvențe de codificare suprapuse. Mutațiile și anomaliile cromozomului sunt responsabile pentru peste 350 de boli, inclusiv cancerul. Deci importanța publicării unei hărți complete a acestui cromozom poate fi cu greu supraestimată.
Tutorial pentru studenti
Curs la universități medicale
Kazan, 2010
Publicat prin decizie a Consiliului Central Coordonator și Metodologic al Universității de Stat de Medicină din Kazan
Compilat de:
Profesor al Departamentului de Biologie Medicală și Genetică
Semenov Valeri Vasilievici,
Asistent al Departamentului de Biologie Medicală și Genetică
Haritonov Vadim Sergheevici
Recenzători:
Universitatea, profesorul Barabanshchikov B.I.,
Șeful Departamentului de Genetică, Statul Kazan
Universitatea Pedagogică, profesorul Zakiev R.K.
Stocarea și implementarea informațiilor ereditare: un manual pentru studenții din anul I ai universităților de medicină / V.V. Semenov, V.S. Kharitonov. - Kazan: KSMU, 2010. - 47 p.
Este destinat studenților primului curs al universităților medicale pentru studiu independent și dobândirea de cunoștințe în domeniul geneticii celulare, mecanismele de funcționare a genelor și implementarea informațiilor conținute în acestea. Manualul acoperă problemele stocării informațiilor genetice și protecția acesteia de factorii mutageni.
© Statul Kazan universitate medicala, 2010 Conţinut
Replicarea ADN-ului…………………………………………………...…………..4
1. Definiție și idei generale despre replicarea ADN-ului…….………….4
2. Perioade de replicare……………….…………………………………5
3. Corectarea (controlul) replicării ………………………………………….12
4. Reduplicarea convariantă ca bază a mutației
variabilitate…………………………………………………………………………….14
5. Metilarea ADN-ului……………………………………………..…………..14
6. Droguri și replicare……..……….……….15
Repararea ADN-ului……….…………………………………………..………......16
1. Ideea generală a reparației…….………………………….…………16
2. Tipuri de reparații……………………………………………………………………… 17
Ideea generală a transcripției………………………………….…..20
1. Perioadele de transcriere………………………………………………………21
2. Erori de împerechere în timpul transcripției. Medicamente
și transcrierea……………………………………………….….…………………………25
3. Procese post-transcripționale. Prelucrare (maturarea ARN)......27
3.1. Interacțiunea pro-i-ARN cu proteina ..…………………………….28
3.2. Metilarea pro-i-ARN…………………………………………………………………………………………………28
3.3. Limitare 5’ – se termină………………..………..……29
3.4. Poliadenilare……………………………………………………………29
3.5. Îmbinarea……………..……………………………..……………….31
3.6. Îmbinare alternativă…………………………………………….32
Difuzare………………………………………………………….…….…...34
1. Ideea generală a emisiunii. suport material
proces…………………………………………………………..………….….…….34
2. Perioade de difuzare…………..……………………………………….……….37
3. Procese post-traduce. Plierea și modificarea proteinelor……….41
Exprimarea genelor. Caracteristicile expresiei genelor la pro- și eucariote…....42
1. Etapele expresiei genelor la eucariote……………………………………………………42
2. Caracteristici ale expresiei genelor la procariote……………………………………….43
3. Reglarea expresiei genelor…………………………………………….……42
3.1. Reglarea expresiei genelor la procariote la nivelul inițierii
transcripții…………………………………….…………....43
3.2. Reglarea expresiei genelor la eucariote…………………..44
REPLICAREA ADN-ului
1. Definiție și idei generale despre replicarea ADN-ului
Replicarea ADN-ului este procesul de transfer de informații de la ADN la ADN ca rezultat al auto-reproducției ambelor catene matrice ale întregii molecule de ADN.
Replicarea este caracteristică ADN-ului și ARN-ului. În prezent, termenul de replicare este cel mai des folosit pentru a se referi la sinteza noilor catene de ADN pe catenele matricei. În această secțiune, ne vom ocupa doar de replicarea ADN-ului.
Din curs şcolar se știe că replicarea la animale are loc în nucleul celular și mitocondrii. În primul rând, molecula de ADN se despiralizează, firele se îndepărtează una de cealaltă și o nouă copie este sintetizată pe fiecare catenă liberă. Sinteza ADN-ului pe catenele se desfășoară în direcții opuse (Fig. 1). După terminarea sintezei, se formează două molecule de ADN. Fiecare moleculă de ADN conține o catenă părinte și o catenă copil, adică. scăpat de naftalină
Orez. 1. Schema de replicare a ADN-ului. Fiecare moleculă de ADN nou sintetizată conține o catenă (linie continuă) a moleculei de ADN șablon, iar cealaltă catenă este nou sintetizată.
jumătate din materialul moleculei de ADN părinte. Prin urmare, o astfel de sinteză se numește semi-conservativă. După încheierea sintezei, două molecule de ADN se îndepărtează una de cealaltă, dar rămân conectate în regiunea centromerului. Moleculele de ADN încep să se îndepărteze complet unele de altele la începutul profazei, când centromerul care le conectează se divide. Toate cele de mai sus sunt incluse în conceptul de sinteză replicativă a ADN-ului. Această sinteză se realizează în perioada S a ciclului celular. Până la sfârșitul acestei perioade, sinteza se oprește. Toate cele de mai sus trebuie să știți, transmițând antrenamentul mai departe curiculumul scolar. În acest manual, vă vom aprofunda înțelegerea replicării, arătând nu numai economia moleculară a acestui proces, ci și semnificația sa medicală.
În primul rând, observăm că oamenii de știință disting un alt tip de sinteză a ADN-ului - reparatorie. Este asociat cu sinteza ADN-ului la locul leziunii sale. Deteriorarea structurii unice a ADN-ului poate apărea în orice etapă a ciclului celular și, datorită pericolului său, trebuie reparată imediat. În caz contrar, celula așteaptă un rând întreg consecințe neplăcute ceea ce a dus în cele din urmă la moartea ei. De exemplu, dacă există extinse
Regenerarea ADN-ului, ei, de regulă, nu se pot recupera și celula moare. Dacă deteriorarea ADN-ului este mică (și apar cel mai adesea), atunci este restabilită destul de repede, sinteza ADN-ului la locul deteriorării este de scurtă durată și nu necesită costuri mari de energie. O astfel de sinteză se numește sinteza ADN neprogramată sau sinteza ADN reparatorie. Dimpotrivă, în timpul replicării, întreaga moleculă de ADN a cromozomului este sintetizată din nou, durata acesteia se măsoară în ore, în timp ce se consumă o cantitate semnificativă de energie și material preparat.
2. Perioade de replicare
Replicarea (precum și pentru orice proces matrice) este caracterizată de trei perioade:
1. Inițiere.
2. Alungirea.
3. Rezilierea.
Iniţiere
Replicarea începe în mai multe puncte ale moleculei de ADN. Acest lucru se datorează faptului că dimensiunea genomului unei celule tipice de mamifer este de aproximativ 109 perechi de baze (bp). ADN-ul unui cromozom uman mediu conține câteva zeci de milioane de perechi de baze. Cu o viteză de replicare de aproximativ 1000 de perechi de baze pe minut va dura câteva zile pentru a finaliza replicarea unei molecule de ADN într-un cromozom. Pentru o celulă, o astfel de perioadă de replicare a genomului este complet inacceptabilă, deoarece în timpul replicării ADN-ului, un proces important, transcripția ARN, nu este efectuat. În timpul replicării ADN-ului, aproape toate genele celulare care îi controlează metabolismul, activitatea vitală, adaptarea și reacțiile celulare specifice sunt dezactivate. Acest lucru se datorează faptului că două procese - sinteza (replicarea) ADN-ului și ARN (transcripția) cu suportul lor enzimatic greoi - nu pot avea loc simultan pe aceeași matrice (ADN matern). Prin urmare, în momentul replicării, transcripția este aproape complet oprită. Acest lucru pune celula într-o stare nefavorabilă, așa cum lipsa expresiei genelor duce la o lipsă a proteinelor enzimatice și, în consecință, la o întrerupere a cursului normal al metabolismului. Cu toate acestea, trebuie remarcat faptul că natura a furnizat mecanisme speciale care reduc riscul de oprire a transcripției pentru celulă. De exemplu, în timpul perioadei de presinteză a ADN-ului (adică înainte de începerea replicării), o cantitate suficientă de toate tipurile de ARN se acumulează în citoplasma celulei în avans pentru a menține sinteza proteinelor în timpul perioadei de replicare a ADN-ului la nivelul necesar pentru viata celulei. Prin urmare, în timpul sintezei ADN-ului, traducerea este efectuată (deși nu în totalitate), în timp ce transcripția practic nu are loc. În orice caz, subliniem încă o dată că timpul în care are loc procesul de replicare a ADN-ului este foarte periculos pentru celulă. Prin urmare, nu este o coincidență că în evoluție
În această perioadă s-au creat mecanisme care reduc la limită timpul de replicare a ADN-ului în cromozomi. Există mai multe astfel de mecanisme, dar unul dintre ele este destul de simplu: replicarea începe simultan în mai multe puncte ale moleculei de ADN și decurge din aceste puncte în direcții diferite. Calculele aproximative au arătat că există câteva mii de astfel de puncte în molecula de ADN a unui cromozom.
Astfel de un numar mare de locurile în care începe replicarea ADN-ului cromozomilor, fără îndoială reduce semnificativ timpul sintezei sale. Punctele în care începe replicarea se numesc punctul de origine al replicării (așa-numitul) sau origine, ori-site (din limba engleză origine - începutul). Aceste puncte sunt mici secțiuni de ADN cu o anumită secvență specifică de nucleotide. Moment de pornire replicarea este atașarea la locurile de origine a proteinelor speciale - proteine inițiatoare.
Să luăm în considerare două prevederi referitoare la ori-site. În primul rând, replicarea nu începe din toate punctele în același timp. În unele puncte, poate trece printr-o secțiune semnificativă, în timp ce în altele poate doar începe. În al doilea rând, de la situsurile ori, procesul de sinteză a ADN-ului se poate deplasa într-o direcție sau în ambele direcții opuse simultan, adică replicarea bidirecțională (Fig. 2). La eucariote, cel mai des se observă al doilea tip de replicare.
Prezența site-urilor ori a condus la crearea conceptului de replicon Un replicon este un fragment al ADN-ului matriceal (matern), pe care are loc sinteza autonomă a ADN-ului fiică. Începe la începutul replicării și se termină în punctul în care enzima încheie replicarea.
A D E DNA
Orez. 2. Schema de localizare a punctelor de origine a replicării. Săgețile indică direcția sintezei ADN-ului din așa-numita.
Repliconul are trei proprietăți principale:
a) este autonom;
b) asigură procesul de sinteză a ADN-ului;
c) are suport enzimatic propriu.
Dacă replicarea este unidirecțională, atunci lungimea repliconului corespunde decalajului dintre site-urile ori, de exemplu, repliconul B - C. Dacă replicarea este bidirecțională (de exemplu, între site-urile ori D - E), atunci doi repliconi G - E și D-F va fi situat între aceste puncte, unde punctul E este sfârșitul a două procese de replicare diferite care au propriul lor provizion enzimatic. După cum au arătat experimentele, este destul de dificil să se determine lungimea medie a unui replicon. În această privință, cel mai adesea un replicon este considerat o secvență de ADN limitată de două situsuri ori.
La originea replicării, apar evenimente care sunt asociate cu atașamentul
unitate la proteina inițiatoare a altor enzime care declanșează o serie de procese pregătitoare. Aceste evenimente sunt enumerate în următoarea ordine:
dar. Formarea unei furci de replicare.
b. Formarea primerului ARN.
dar. Formarea unei furci de replicare
Formarea unei furci de replicare are loc la originea replicării. Furculița de replicare sunt catene libere de ADN șablon care s-au îndepărtat unele de altele, pe care are loc sinteza catenelor ADN fiice. Figura 3 prezintă formarea furcilor de replicare.Două furci de replicare formează un ochi de replicare. În ambele furci de replicare, sinteza ADN-ului are loc în direcții opuse. În ochiul replicării, furcile se mișcă, derulând ADN-ul, în direcții opuse.
Formarea furcilor de replicare este precedată de o serie de procese pregătitoare, care, de regulă, sunt aranjate în următoarea ordine:
Eliberarea moleculei de ADN de la legarea de histone din nucleozom.
Despiralizarea ADN-ului.
ruperea legăturilor de hidrogen dintre catenele ADN-ului matricei.
Divergența catenelor de ADN.
Fixarea catenelor matricei ADN.
Eliberarea moleculei de ADN din legătura sa cu histonele este un element necesar al perioadei de inițiere. În nucleul celulelor, ADN-ul nu este aproape niciodată în stare liberă. Este asociat cu proteinele tetramer histonă și histona H1. În plus, diverse proteine non-histone, metale divalente, lipide și alți compuși interacționează cu ADN-ul. Toate acestea conduc la o puternică spiralizare a secțiunilor individuale de ADN în perioada pre-sintetică.
Cum este eliberat ADN-ul din legăturile cu moleculele din jur nu este complet clar. Cu toate acestea, se știe că acest proces se bazează pe mai multe reacții chimice efectuate de enzime - metilarea, acetilarea și fosforilarea grupurilor încărcate pozitiv de proteine histone și nonhistone. Ca urmare, proteinele își pierd sarcina pozitivă, legăturile lor cu molecula de ADN încărcată negativ devin slabe. Aceasta promovează eliberarea de ADN.
Imediat după eliberarea din legăturile cu proteina (sau în paralel cu aceasta), are loc procesul de despiralizare a ADN-ului. Acest proces este necesar pentru eliberarea ulterioară a catenelor de ADN din legarea între ele. Un alt proces este, de asemenea, important. Catenele moleculei de ADN nu sunt doar paralele între ele, ci sunt răsucite reciproc (vezi Fig. 4).
Există multe moduri de a derula spiralele. Cele mai frecvent utilizate metode sunt prezentate în Figura 5. În mod evident, metoda A nu este în întregime convenabilă, deoarece când un capăt este fixat și capetele opuse sunt separate, se creează forțe suplimentare, ducând la formarea de superbobine secundare. Cu a doua metodă (capătul opus este gratuit)
desfășurarea va face ca capătul liber să se rotească, iar acest lucru poate transforma nucleul într-un amestecător submolecular, ceea ce nu este sigur pentru celulă.

Furcă stânga Furcă dreapta
Două furci de replicare
 Orez. 3. Schema formării furcilor de replicare la punctul de origine a replicării. Săgețile punctate arată direcția de mișcare a furcilor
Orez. 3. Schema formării furcilor de replicare la punctul de origine a replicării. Săgețile punctate arată direcția de mișcare a furcilor
Orez. 4. Schema poziție relativă catene de ADN. Legăturile de hidrogen între fire nu sunt prezentate.
Pentru derularea lor se folosesc enzime care taie, rotesc si reunesc firele.
Principiul derulării helixului ADN nu este complet clar. Una dintre opțiunile posibile este prezentată în Figura 6. Esența sa este că o enzimă specială produce o rupere temporară a unei singure catete într-o catenă de ADN. Și prin această tăietură trece (este târât) șirul opus de ADN. În acest caz, o tură a spiralei se îndreaptă. Incizia este apoi închisă. Proces
tăierea, desfășurarea și cusătura pot fi repetate de multe ori pe măsură ce furculița de replicare se mișcă de-a lungul moleculei de ADN.
Toate reacțiile asociate cu desfășurarea catenelor de ADN sunt efectuate de enzimele topoizomerazei. Este oportun să rețineți că aceste enzime acționează într-un mod similar într-un alt proces - transcripția. Trebuie remarcat faptul că, în același timp, enzimele helicaze speciale rup legăturile de hidrogen dintre catenele de ADN opuse (opuse), făcându-le libere.
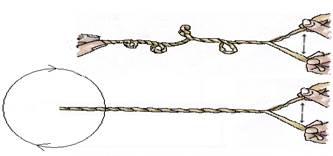
Orez. 5. Modalități de a desfășura frânghia. A - capătul opus este fix, B - capătul opus este liber
Catenele de ADN eliberate din duplex nu pot rămâne în stare liberă mult timp. Ele fie se recombină, fie interacționează cu reactivii activi din spațiul înconjurător, fie formează ace de păr. Oricare dintre opțiuni poate perturba cursul normal al replicării. Pentru a preveni acest lucru, există proteine speciale - proteine SSB. Aceste proteine au o afinitate pentru ADN-ul monocatenar și, odată cu apariția acestuia din urmă, se conectează imediat cu acesta pe toată lungimea sa, formând un cadru rigid, prevenind nu numai reacțiile sale, ci și formarea acelor de păr (Fig. 7, 1).
O proprietate importantă a acestor proteine este că ele lasă bazele nucleice ale unei catene de ADN monocatenar deschise pentru reacții, ceea ce este extrem de important pentru sinteza ulterioară a catenelor ADN fiice.
b. Formarea primerului ARN
După finalizarea tuturor proceselor de mai sus, furca de replicare este aproape gata pentru sinteza ADN-ului și arată așa cum se arată în Figura 7, 1. Cu toate acestea, un detaliu esențial lipsește pentru pregătirea completă a ADN-ului pentru replicare. Sinteza catenelor fiice în conformitate cu catenele de ADN fără șablon este realizată de enzima ADN polimeraza. Cu toate acestea, această enzimă nu poate începe procesul de sinteză a unui fir nou. El este capabil doar să continue sinteza unui lanț deja început. Prin urmare, o altă enzimă, ARN polimeraza dependentă de ADN (ARN primaza), mai întâi sintetizează o mică porțiune de ARN pe ambele catene ale matricei ADN care au divergit în furculița de replicare. Acest ARN se numește primer ARN (primer ARN sau pur și simplu primer) (în Fig. 7 sunt marcați cu literele „A” și „B”). Primerii ARN sunt sintetizați în paralel cu catenele șablon de ADN conform principiului complementarității. Mai mult, pe o catenă, sămânță de ARN
(în Fig. 7 este desemnat ca „A”) este sintetizat, pornind de la locul ori spre furculița de replicare, pe catena opusă, primerul ARN (în Fig. 7 este desemnat „B”) este sintetizat, plecand de la site-ul ori din partea lui.
Tăiați firul nr. 2 și trageți prin tăierea firului nr. 1


Orez. 6. Schema despiralizarii unui segment al unei molecule de ADN prin taierea unui fir, trasarea altuia, derularea si cusatura taieturii dupa derularea sectiunii. Legăturile de hidrogen între fire nu sunt prezentate.
Sinteza a doi primeri ARN pe două catene opuse de ADN matriță încheie perioada de inițiere.
Elongaţie
Enzima ADN polimerază începe sinteza noilor catene de ADN de la capetele 3’ ale a doi primeri ARN (A și B în Fig. 7, p. 2). Cu o astfel de sinteză, moleculele de ADN fiice de pe catenele matricei vor fi sintetizate în direcții opuse.
Alungirea începe cu adăugarea unei nucleotide la capătul 3’ al primerului ARN. Această atașare este realizată de enzima ADN polimerază.ADN polimeraza atașează a doua, a treia etc.la prima nucleotidă. nucleotidă în conformitate cu principiul complementarității cu nucleotidele situate pe catena matricei ADN. Bazele nucleotidelor noii catene și ale matricei sunt legate între ele prin legături de hidrogen. Deoarece sinteza unei noi catene de ADN începe de la capătul 3’ al primerului de ARN, de care este atașat capătul 5’ al primei nucleotide ADN, se obișnuiește să spunem că sinteza catenei de ADN fiice are loc în direcția 5’-3’. Pe acel fir de matrice unde sinteza
al catenei fiice se îndreaptă spre furca de replicare, merge continuu, iar pe măsură ce catena se desfășoară, fragmentul de ADN sintetizat se va lungi constant. Cu cât furculița de replicare avansează mai mult, cu atât catena de ADN nou sintetizată va fi mai lungă. Această catenă de ADN se numește continuu, conducător sau conducător. Ulterior, nu se formează primeri ARN pe catena de ADN șablon, din care este replicată catena principală. Mai dificilă este sinteza ADN-ului fiice pe catena opusă a ADN-ului matern, care are o direcție de 5 '-3'. La primerul ARN sintetizat pe această catenă (indicat ca „B” în Fig. 7), nucleotidele vor fi, de asemenea, atașate la capătul 3’, iar o nouă catenă va fi sintetizată departe de furcul de replicare (Fig. 7, 2). ). Această catenă de ADN fiică se numește rămas în urmă. Astfel, în furca de replicare sunt sintetizate simultan două catene fiice de ADN - cea de conducere (este sintetizată în direcția mișcării furcii de replicare) și cea mai întârziată (este sintetizată în sens opus mișcării furcii de replicare), adică direcția de sinteză a ambelor lanțuri fiice este opusă.
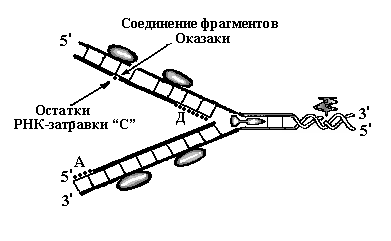
Sinteza catenei întârziate de ADN nu are loc în mod constant, ci în fragmente. După terminarea sintezei unui fragment în apropierea furculiței de replicare, primerul ARN „C” este sintetizat din nou. De la capătul 3’ al acestui primer, sinteza catenei fiice de ADN începe din nou în direcția primerului ARN „B”, iar când este atins, sinteza se oprește din nou - s-a format un al doilea fragment de ADN, care începe de la primerul ARN „C”. De fapt, pe firul întârziat, avem două fragmente, care constau din ARN și ADN conectate între ele. Un fragment de ADN începe cu primerul ARN „B”, al doilea - cu primerul ARN „C”. Aceste fragmente de ADN sunt numite fragmente Okazaki (Fig. 7.3).
Rundele ulterioare de replicare se repetă - topoizomeraza desfășoară o altă rotație a helixului ADN, helicaza rupe legăturile de hidrogen dintre catenele de ADN și acestea diverg, proteinele SSB fixează catenele, catena principală continuă să se lungească, al treilea fragment al primerului ARN este sintetizat. pe firul întârziat - „D” și cu capătul său 3 ', începe să fie sintetizat un nou fragment Okazaki. Apoi, pe catena de ADN rămasă, primerii de ARN sunt distruși și fragmentele de ADN rămase sunt combinate într-un singur lanț (Fig. 8). Astfel, 4 procese se desfășoară continuu pe lanțul întârziat: formarea de noi primeri ARN, sinteza fragmentelor Okazaki de la capătul lor 3’, distrugerea primerilor ARN și reunificarea fragmentelor într-un singur lanț.
Încetarea
Sinteza ADN-ului continuă până când două furculițe de replicare se întâlnesc sau până când furculița de replicare ajunge la capătul moleculei de ADN de pe cromozom. După ce furcile de replicare se întâlnesc, catenele de ADN fiice sintetizate din furcile adiacente sunt conectate printr-o enzimă. Terminarea este mai dificilă dacă furculița de replicare a ajuns la capătul moleculei de ADN. Dar și aici, enzimele speciale opresc replicarea (Fig. 9).
3. Corectarea (controlul) replicării
În timpul replicării ADN-ului are loc un proces important de corectare a nucleotidelor inserate (Fig. 10). Toate cele patru nucleotide intră în regiunea sintezei ADN-ului (în Fig. 10 sunt desemnate ca „nucleotide candidate”). ADN polimeraza selectează precursorul dorit prin împerecherea de probă cu o nucleotidă matriță ADN.

Orez. 7. Modele de replicare a ADN-ului
 Corectitudinea nucleotidei selectate este evaluată prin mai mulți parametri, dintre care doi sunt cei mai semnificativi. În primul rând, nucleotida selectată trebuie să formeze tot atâtea legături de hidrogen câte nucleotide de pe catena șablon.
Corectitudinea nucleotidei selectate este evaluată prin mai mulți parametri, dintre care doi sunt cei mai semnificativi. În primul rând, nucleotida selectată trebuie să formeze tot atâtea legături de hidrogen câte nucleotide de pe catena șablon.
Orez. 8. Schema de replicare a ADN-ului - etapa de alungire. Pe catenă întârziată, a fost sintetizată a treia regiune a primerului ARN, „D”. Primerii ARN „B” și „C” sunt distruși și fragmentele Okazaki sintetizate de la capetele lor sunt conectate într-un singur fir. Pe lanțul principal, sinteza este continuă, primerul ARN „A” este distrus
Origins Leading Chain
Lant întârziat
Replicon
Mișcarea furcii Mișcarea furcii
Orez. 9. Schema sintezei ADN-ului în două furcuri de replicare
În al doilea rând, distanța dintre schelele zahăr-fosfat trebuie să fie constantă și să corespundă la trei inele heterociclice în două baze (un inel corespunzător unei baze pirimidinice într-o catenă și două inele corespunzând unei baze purinice în cealaltă catenă). Dacă nucleotida candidată și nucleotida din matrice conțin două inele în moleculă sau câte unul, atunci conexiunea nu are loc.
Verificarea includerii corecte a nucleotidei în lanțul de creștere se efectuează de două ori: o dată înainte de includerea acesteia în lanțul de creștere și a doua oară înainte de includerea următoarei nucleotide.
După ce testul de compatibilitate a bazelor nucleice a trecut, se formează o legătură în coloana vertebrală zahăr-fosfat.
4. Reduplicarea convariantă ca bază a variabilității mutaționale
Acest termen a fost introdus în uz științific de către geneticianul Timofeev-Resovsky. Literal, reduplicarea convariantă înseamnă auto-reproducere cu schimbare. Acest proces stă la baza întregului proces evolutiv.
În ciuda acurateței ridicate a proceselor de replicare și a unui sistem de corecție eficient, catenele de ADN nou sintetizate conțin întotdeauna perturbări. Aceste defecte sunt denumite cel mai frecvent „mutații genetice”. Calculele au arătat că acestea apar cu o frecvență de 1 eroare la 10 10 perechi de baze. Reduplicarea care apare cu erori se numește reduplicare convariantă.
Încălcările se bazează pe o varietate de motive. De exemplu, o concentrație mare sau invers foarte scăzută de nucleotide la locul de sinteză, pierderea spontană a bazelor purinice (vezi mai jos), dezaminarea citozinei, care este transformată în uracil sub acțiunea radiațiilor ultraviolete, prezența mutagenilor chimici la locul sintezei etc. Soarta erorilor rezultate este ambiguă. Ele pot fi corectate prin procese de reparare (vezi mai jos). Aceasta este cea mai favorabilă opțiune pentru celulă. În unele cazuri, deteriorarea moleculei de ADN nu este reparată. Există două scenarii posibile pentru desfășurarea evenimentelor. Dacă deteriorarea unei molecule de ADN a afectat o regiune funcțional inactivă a ADN-ului, atunci din punct de vedere fenotipic o astfel de eroare nu va fi detectată și, de regulă, nu vor fi observate consecințe grave. Este o problemă diferită dacă eroarea de împerechere a avut loc în secvențele de nucleotide ale oricărei gene. În acest caz, probabilitatea de apariție a tulburărilor fenotipice crește. Acest lucru poate duce la moartea celulei sau a întregului organism. În acest caz, împreună cu celula moartă, se elimină și defectul ADN. În cele din urmă, ar trebui să se țină seama de faptul că mutațiile genice stau la baza variabilității mutaționale. Și duce la apariția mai multor alele, care fac fondul genetic al populațiilor mai plastic.
5. Metilarea ADN-ului
Și în concluzie, este necesar să ne oprim asupra unui proces important care are loc în momentul sau imediat după sinteza catenelor fiice de ADN - metilarea catenelor nou sintetizate. La om, o enzimă specială, ADN-metilaza, metilează citozina prin adăugarea de grupări CH3. Procesul de metilare este cel mai probabil necesar pentru formarea structurii cromozomilor (ambalarea ADN) și reglarea transcripției genelor. În plus, metilarea contribuie la inactivarea unui cromozom X la mamifere. La bacterii, metilarea protejează ADN-ul de a fi tăiat de propriile enzime.
coloana vertebrală a fosfatului de zahăr
Hidrogen
| | |||||
lanț pentru copii
| | |||||
| |
|||||||||
Șablon de ADN T
 |
||||||||
Nucleotide Challenger
Orez. 10. Schemă de selectare a unei baze complementare la o nucleotidă de citozină (C). Dintre candidați, doar nucleotida guanină (G) este compatibilă cu această nucleotidă; formează trei legături de hidrogen și are două inele
6. Droguri și replicare
Procesul de replicare este afectat de o varietate de factori și, în special, de medicamente. De exemplu, daunomicina și alte medicamente antitumorale au o structură ciclică plată în molecula lor, care este construită (intercalată) între perechile de baze. Acest lucru duce la o modificare locală a structurii ADN-ului, ca urmare a căreia enzimele de replicare nu mai funcționează. De exemplu, topoizomeraza își pierde capacitatea de a despira ADN-ul la locul de intercalare. O astfel de alchilare
Substanțe precum tiofosfamida și altele modifică bazele prin adăugarea de grupări alchil la acestea. Dacă enzimele de replicare întâlnesc această bază, activitatea lor se poate opri.
Inhibitorul ADN-topoizomerazei - novobiocina, interferând cu activitatea enzimei, oprește despiralizarea ADN-ului și, în consecință, sinteza ARN-ului.
Rezumat. Replicarea ADN-ului este procesul de transfer precis al informațiilor de la ADN la ADN ca rezultat al auto-reproducției moleculelor matricei. Există trei perioade în replicare.
1. Iniţiere. Se formează o furcă de replicare și se formează un primer ARN. Sinteza începe simultan la mai multe regiuni ADN. Înainte de sinteză, ADN-ul este despiralizat, legăturile de hidrogen sunt rupte și firele se îndepărtează unele de altele.
2. Elongaţie. Sinteza ARN începe cu un primer ARN și se desfășoară simultan pe ambele catene de ADN matern. Pe o catenă, sinteza se desfășoară continuu, pe cealaltă - în fragmente (fragmente Okazaki).
3. Încetarea. Sinteza ARN se termină la întâlnirea furcilor de replicare sau la capătul moleculei de ADN.
| REPARAREA ADN-ului |
1. Idee generală de reparație
Procesul de reparare este caracteristic tuturor ființelor vii. Aceasta este una dintre principalele proprietăți ale materialului ereditar, direct legată de intensitatea proceselor de îmbătrânire, dinamica dezvoltării bolilor tumorale, formarea malformațiilor congenitale, apariția patologiei ereditare etc. Modificări ADN care nu au suferit reparații. au jucat (și joacă) un rol important în formarea evolutivă a noilor proprietăți adaptative ale celor vii.
Să ne oprim asupra a trei principii importante caracteristice procesului de reparare.
1. Enzimele de reparare determină locația celor mai multe daune pe molecula de ADN prin denaturarea sa locală la locul leziunii.
Eficacitatea oricărui sistem de reparare, în primul rând, depinde de cât de repede și de exact enzimele detectează locurile de deteriorare a ADN-ului. Întrucât natura acestora din urmă este extrem de diversă, este greu de imaginat existența a numeroase enzime care recunosc „lor” daune specifice ADN-ului. De fapt, totul s-a dovedit a fi mai ușor. Orice încălcare a structurii primare a ADN-ului duce la aceleași consecințe - imediat apare o încălcare în structura celui de-al doilea, al treilea etc. comanda, adica denaturarea locală a ADN-ului are loc la locul leziunii. Zonele de denaturare care au apărut în ADN (spre deosebire de deteriorarea primară) nu sunt specifice. Acest lucru permite enzimelor speciale să detecteze rapid denaturate
secţiuni ale ADN-ului şi conectează sisteme de reparaţii specifice pentru restaurarea acestora.
2. Secțiunea deteriorată de pe o catenă de ADN este restaurată în conformitate cu secțiunea opusă intactă a celei de-a doua catene. Când două fire sunt deteriorate, firele surori ale cromatidelor sunt implicate în repararea daunelor.
S-a observat o regularitate că atunci când ADN-ul este expus la factori mutageni, deteriorarea se referă în principal la o catenă de ADN. Deteriorarea simultană a ambelor catene nu are loc des și, de obicei, se termină cu moartea celulară. Restaurarea unei zone deteriorate nu este dificilă dacă există un plan pentru structura sa originală (înainte de deteriorare). Un astfel de plan este întotdeauna disponibil - aceasta este secvența de nucleotide de pe a doua catenă de ADN opusă, adică. în catena de ADN opusă locului leziunii. Aici sunt localizate nucleotidele care reflectă în mod complementar secvența de nucleotide a zonei deteriorate. Prin urmare, multe sisteme de reparații „repară” zona deteriorată prin construirea acesteia cu nucleotide complementare, în conformitate cu planul zonei opuse.
Este o problemă diferită dacă ambele catene de ADN sunt deteriorate în același timp. Acest lucru se întâmplă atunci când o moleculă de ADN cu deteriorare intră într-o perioadă de sinteză a ADN-ului. În acest caz, sinteza catenelor fiice de ADN are loc pe ADN-ul defect, iar o catenă fiică defectă este sintetizată pe catena deteriorată. Dar chiar și în acest caz, planul zonelor deteriorate este păstrat în molecula de ADN, care s-a format ca urmare a replicării unei catene de ADN intacte. Cu convergența maximă a cromatidelor surori în profaza meiozei și mitozei, repararea catenelor de ADN deteriorate are loc în conformitate cu secțiunile intacte ale moleculei de ADN ale cromatidei surori.
În organismele diploide, la repararea unei regiuni ADN deteriorate, este posibil să se utilizeze planul structural al unei regiuni ADN nedeteriorate a unui cromozom omolog.
3. Energia pentru procesul de reparare este extrasă în principal din ATP. Dar, în unele cazuri, se folosește energia unui foton de lumină, care activează enzimele de reparare.
Repararea ADN-ului este un proces dependent de energie. Compușii cu înaltă energie (de exemplu, ATP) sunt utilizați ca furnizor de energie. Cu toate acestea, există o excepție - aceasta este fotoreactivarea (repararea luminii). Enzima care realizează această reacție folosește energie lumina soareluiși nu se acumulează în legăturile chimice ale ATP. Această enzimă se numește fotoliază. Enzima repară daunele asociate cu formarea de dimeri de timină (vezi mai jos).
2. Tipuri de reparații
În primul rând, toate tipurile de reparații (sau reacții) sunt împărțite în două mari grupuri - directe și indirecte. Reacțiile directe sunt simple și inconsistente.
coloana vertebrală a fosfatului de zahăr
 UV T A
UV T A
 |
|||
 |
|||
| |
|||
Orez. 11. Schema formării dimerilor de timină sub acţiunea iradierii UV. Legăturile chimice formate între timinele unui fir sunt prezentate prin linii punctate
înseamnă acţiune asupra obiectului deteriorat. Nu se efectuează „lucrare preliminară”. Aproape toate funcționează până în perioada de sinteză a ADN-ului. Acestea includ fotoreactivarea. Cu un tip de reparație indirectă, deteriorarea ADN-ului existentă este mai întâi excizată. Ca urmare, se formează un gol pe una dintre catene, care este construit de enzime speciale în conformitate cu planul de aranjare a nucleotidelor pe catena opusă. Acest tip de reparație include repararea exciziei și o serie de altele.
Tip reparatie directa
Fotoreactivare
Este un fapt incontestabil că lumina ultravioletă (UV) dăunează moleculei unice de ADN. Daunele se referă în principal la baze pirimidinice - timină și citozină. Sub influența luminii UV,
redistribuirea valențelor în baze pirimidinice, i.e. între
două timine, două citozine sau timină și citozină una lângă alta pe o singură șuviță. Consecința acestui lucru este apariția unei legături chimice neobișnuite între baze și apariția unui dimer (două nucleotide) - timină (timină-timină), citozină (citozină-citozină) sau timină-citozină. Legătura de hidrogen dintre nucleotidele opuse este distrusă în acest caz (Fig. 11). Există dovezi că dimerii de pirimidină sunt capabili să activeze procese care conduc la dezvoltarea bolilor tumorale.
Mutația rezultată este reparată de mai multe sisteme de reparare. Una dintre ele este fotoreactivarea. Principala enzimă a acestei reacții, proteina fotoliază, are o structură proteică complexă. Într-o parte a moleculei există un centru sensibil la lumină care percepe fotonii luminii albastre și activează enzima. Enzima în această stare găsește dimeri în molecula de ADN, rupe legăturile UV formate între timine și restabilește legăturile de hidrogen între catenele pirimidină-purină. La sfârșitul ciclului, enzima se îndepărtează de ADN.
Trebuie remarcat faptul că, în acest tip de reparații, enzima acționează direct asupra daunei, restabilind-o. Acesta este un tip clasic de reparație directă.
Tip indirect de reparație
Reparație de excizie
Incizie Lărgirea golului Închiderea golului Cusătură
(Endonuclează) (Exonuclează) (ADN polimerază) (Ligază)

dimer de timină
Orez. 12. Schema reparației prin excizie a dimerilor de timină
Este posibil ca dimerii de timină să nu fie reduceți de sistemul de fotoreactivare din mai multe motive. În acest caz, enzimele sistemului de reparare excizională sunt activate. Acest tip de reparație face posibilă corectarea a până la 95% din mutațiile spontane. Există mai multe tipuri de reacții de reparare prin excizie. Ei sunt uniți de faptul că baza deteriorată, nucleotida sau baza împerecheată incorect (nepotrivire) este mai întâi tăiată complet din lanț, iar apoi este introdusă o nouă nucleotidă (sau nucleotide) care este complementară cu nucleotida lanțului opus. Să luăm ca exemplu o reparație prin excizie
cation al nucleotidelor (dimeri de timină). Reacția constă din mai multe etape. În prima etapă, enzima endonuclează găsește catena de ADN deteriorată și face o incizie la locul leziunii. O altă enzimă, exonucleaza, lărgește crestătura prin „mușcarea” nucleotidelor de la ambele capete ale crestăturii împreună cu dimerul de timină. Decalajul rezultat este construit de la un capăt al catenei de ADN de o enzimă specială, ADN polimeraza. În acest caz, nucleotidele inserate sunt complementare cu nucleotidele catenei intacte de ADN. Un mic gol rămâne neumplut, care este cusut împreună de ADN ligază (Fig. 12).
INTRODUCEREA GENERALĂ A TRANSSCRIPȚIEI
Expresia genelor este realizarea de informații genetice codificate în ADN prin transcrierea și traducerea acestuia. Transcrierea este primul pas în exprimarea genelor. Constă în traducerea informațiilor conținute în genă în ARN prin sintetizarea acestuia din urmă pe o catenă de ADN a genei. Ca urmare a transcripției, se sintetizează toate tipurile de ARN - informațional (ARNm), ribozomal (ARNr), transport (ARNt) și altele (regulator, nuclear mic etc.). Toți iau parte la expresia genelor, dar numai ARNm transferă informații despre structura proteinei din textul nucleotid al ADN-ului către textul aminoacizi al proteinei. Toate celelalte tipuri de ARN asigură implementarea eficientă a acestui proces.
Esența transcripției este următoarea: enzime speciale pregătesc molecula de ADN în regiunea genei pentru transcripție (desfășoară helixul ADN-ului, rup legăturile de hidrogen dintre catene etc.). Enzima ARN polimeraza dependentă de ADN sintetizează ARN-ul pe catena matriță de ADN de la punctul de început până la punctul final al transcripției. Sinteza ARN-ului constă în acumularea secvenţială de nucleotide în el care sunt complementare catenei matriţe a ADN-ului. Excepția este ARN-ul uracil, care este inserat împotriva adeninei catenei șablon de ADN.
Multe enzime iau parte la transcripție, dar sinteza ARN este realizată direct de enzima ARN polimeraza dependentă de ADN (sau pur și simplu ARN polimeraza). La procariote, toate tipurile de ARN (ARNm, ARNr, ARNt) sunt sintetizate de o ARN polimerază, în timp ce la eucariote sunt sintetizate de trei enzime diferite: ARN polimeraza I, ARN polimeraza II și ARN polimeraza III.
ARN-urile mesager la eucariote sunt transcrise prin ARN polimeraza II.
Transcripția este un proces complex în mai multe etape și ARN polimeraza singură nu este capabilă să o furnizeze pe deplin. În diferite etape ale transcripției, diferite subunități proteice sunt atașate la ARN polimerază și, dimpotrivă, scindate, ceea ce îi modifică activitatea în conformitate cu cerințele acestei etape.
Transcripția, ca toate procesele de sinteză a șablonului la pro- și eucariote, constă din trei etape: inițiere, alungire și terminare. În cele ce urmează, procesul de transcripție va fi luat în considerare numai la eucariote.
1. Perioade de transcriere
Iniţiere
Inițierea este o etapă pregătitoare. În această etapă, au loc o mulțime de procese sincronizate în timp. Să luăm în considerare două dintre ele:
1. Formarea complexului iniţiatic.
2. Formarea unui „ochi transcripțional”.
complex inițiator
promotor
Orez. 13. Atașarea ARN polimerazei și transcripția
factori la promotorul cutiei TATA la eucariote
Formarea complexului inițiator are loc la promotor. ARN polimeraza însăși, de regulă, nu se poate lega de promotor. Prin urmare, o proteină specifică interacționează mai întâi cu o regiune specifică de pe promotor. Această regiune conține o secvență specifică de nucleotide. Această secvență se numește cutia Pribnov. . Această secvență de nucleotide este bogată în nucleotide de adenină și timină - TATA. De aici și celălalt nume pentru această regiune a promotorului
Pagina 1
Informațiile genetice sunt criptate în molecula de ADN folosind un cod pe care noi îl numim cu modestie universal. Aceasta înseamnă că, dacă se știe cum se produce transmiterea informațiilor ereditare într-o singură celulă, atunci se cunoaște și mecanismul molecular al acestei transmiteri în orice alte celule, și nu numai al aceluiași organ sau organism, ci în general la toate ființele vii. .
Informațiile genetice sunt înregistrate ca perechi de baze pe dubla helix ADN. Un bloc de informații care corespunde unei proprietăți sau trăsături este denumit genă. Setul complet de gene se numește genom. Genomul bacterian include câteva mii de gene dispuse liniar pe o macromoleculă de ADN numită cromozom. La eucariote, firele de ADN sunt formațiuni complexe, numite și cromozomi, dintre care pot fi multe per celulă. Cromozomii eucarioți sunt separați de citoplasmă printr-o membrană nucleară, formând nucleul. Diferența citologică în structura aparatului genetic este principala în diferența dintre procariote și eucariote, restul caracteristicilor sunt fie suplimentare, fie corelate. În plus față de cromozom, molecule circulare mici de ADN numite plasmide pot fi prezente în procariote, care nu sunt necesare pentru genomul unei anumite specii. Plasmidele variază ca mărime și număr în celulă de la una la mai multe zeci. Plasmidele mari pot conține sute de gene. Plasmidele sunt purtători extracromozomiali de informație genetică.
Informația genetică este codificată în secvența de baze de-a lungul lanțului ADN. Cu fiecare diviziune celulară, această informație este copiată și transferată în celula fiică. În plus, pe baza acestor informații, proteinele corespunzătoare sunt sintetizate în celule. Fiecare genă din molecula de ADN conține instrucțiunile necesare pentru sinteza unei proteine.
Informațiile genetice despre structura oricărei specii biologice sunt stocate în siguranță și nu pot fi pierdute atâta timp cât există un număr suficient de indivizi pentru a reproduce specia. Totuși, poluarea din ce în ce mai mare a biosferei cu deșeuri antropice duce la modificări ireversibile ale programului genetic uman.
Informația genetică este stocată ca o secvență de nucleotide într-o moleculă de ADN liniară. ADN-ul poate fi împărțit în secțiuni continue (gene), fiecare dintre ele conține secvența de aminoacizi a unei proteine. Genele sunt separate prin regiuni reglatoare de care se leagă ARN polimerazele și proteinele represoare.
Informația genetică este aparent încorporată în secvența a patru nucleotide (baze) care formează lanțul polinucleotidic al ADN-ului.
Informația genetică este transmisă de la părinte la fiică prin replicarea ADN-ului (sinteză) - Informația genetică este stocată în ADN până când este nevoie și apoi convertită în instrucțiuni pentru sinteza proteinelor unei secvențe specifice în timpul transcripției. Instrucțiunea genetică este copiată pe molecula de ARN polimeric. La rândul său, interacționează cu ARN-ul aminoacil-t specific corespunzător, rezultând o adăugare secvențială de aminoacizi.
Informațiile genetice din toate ființele vii sunt codificate în moleculele de ADN. Genomul uman conține aproximativ 3x109 nucleotide sub formă de 100.000 de gene.
Informația genetică necesară controlului sintezei proteinelor cu o structură strict definită este codificată de secvența de nucleotide a lanțului de ADN.
Informația genetică, totuși, poate fi folosită nu numai rațional, ci și irațional.
Informația genetică controlează o persoană la nivel molecular și biologic în mod automat, fără participarea conștiinței noastre. Într-adevăr, nu ne gândim la modul în care funcționează inima, plămânii, ficatul, stomacul, vezica urinară.
Informația genetică a bacteriilor nu se limitează la ADN-ul situat în nucleoidul celulei bacteriene. După cum sa menționat deja în secțiunile anterioare ale cărții, elementele extracromozomiale, numite colectiv plasmide, servesc și ca purtători de proprietăți ereditare. Spre deosebire de ADN-ul echivalentelor nucleare, nucleoizii, care sunt organitele unei celule bacteriene, plasmidele sunt elemente genetice independente.
| Reprezentare schematică a celor patru baze nucleotidice care alcătuiesc molecula de ADN.A-adeiina. T - timină. G - gua-nin. C - citozina.| Nucleotidele se combină cu grupele de zahăr C, care la rândul lor sunt legate între ele prin grupări fosfat F, formând un lanț. Întreaga moleculă de ADN constă dintr-o catenă dublă sub formă de helix. Două lanțuri simple sunt conectate prin legături de hidrogen între grupările adenină și timină sau între grupările guanină și citozină. |













