Caracteristicile structurale ale țesutului muscular ale funcțiilor îndeplinite. Structura și proprietățile de bază ale țesutului muscular.
Capitolul 9
Capitolul 9
tesut muscular (textus muscularis) numite țesuturi care sunt diferite ca structură și origine, dar similare ca capacitate de contractare. Ele asigură mișcarea în spațiul corpului în ansamblu, părțile acestuia și mișcarea organelor din interiorul corpului (inima, limba, intestinele etc.).
Condițiile de sacrificare și metoda de prelucrare după sacrificare afectează direct calitatea cărnii. Înainte de sacrificare, animalele sunt supuse unei operații de asomare. Uimirea se poate face în mai multe moduri și anularea sensibilității animalului la durere, oprirea centrilor nervoși ai relațiilor de viață, menținând în același timp funcționalitatea bulbului inimii, pentru a înjunghia complet după sângerare.
Stabilirea corecta a operatiunilor de urmarire de injunghiere, sangerare, depilare, decojire, indepartare a capului si membrelor, picioarelor, eviscerarea si decaparea carcaselor afecteaza favorabil aspectul.Carcasele comerciale ajuta la reducerea infectiei microbiene si asigura o buna stabilitate la depozitarea carnii.
Capacitatea de a se contracta cu o schimbare a formei este deținută de celulele multor țesuturi, dar în țesuturile musculare această abilitate devine principala funcție.
9.1. CARACTERISTICI MORFOFUNCȚIONALE GENERALE ȘI CLASIFICARE
Principal caracteristici morfologice elemente ale țesutului muscular - o formă alungită, prezența miofibrilelor și miofilamentelor dispuse longitudinal - organele speciale care asigură contractilitatea, localizarea mitocondriilor lângă elementele contractile, prezența incluziunilor de glicogen, lipide și mioglobină.
Transformarea după sacrificii și efectul lor asupra atributelor cărnii. După sacrificarea animalelor, în carne au loc o serie de modificări, care provoacă schimbari importanteîn proprietăţile sale organoleptice şi tehnologice. Aceste transformări pot fi normale, cum ar fi rigiditatea musculară, maturizarea, febra sau anomalii, cum ar fi căldura și putrefacția. Transformările normale îmbunătățesc valoarea nutritivă a cărnii.
Rigiditatea musculară apare oarecum din sacrificarea animalului. Ca urmare a acestui proces, mușchii își pierd elasticitatea și devin rigidi sub acțiunea acidului lactic, care se coagulează. proteineși provoacă contracția musculară. Apariția rigidității și durata acesteia sunt determinate de mai mulți factori: temperatură mediu inconjurator, specie, vârstă, stare fiziologică și oboseală, sănătatea animală etc. perioada de rigiditate durează de obicei aproximativ 24 de ore urmată de maturare.
Organele contractile speciale - miofilamente sau miofibrile, asigură contracția care are loc atunci când cele două proteine fibrilare principale - actina și miozina - interacționează în ele, cu participarea obligatorie a ionilor de calciu. Mitocondriile furnizează energie pentru aceste procese. Furnizarea surselor de energie este formată din glicogen și lipide. Mioglobina este o proteină care asigură legarea oxigenului și crearea rezervei sale în momentul contracției musculare, când vasele de sânge sunt comprimate (aportul de oxigen este redus brusc).
Maturarea. În procesul de maturare, cantitatea de acid lactic scade și au loc procesele de hidroliză a substanțelor proteice. Ca urmare a acestor modificari, carnea capata o textura moale, devine suculenta si suculenta, cu un gust foarte placut, iar culoarea rosie devine mai putin intensa datorita combinarii pigmentilor cu oxigenul. Cu cât temperatura este mai mare, cu atât procesul de coacere este mai lung. Lungimea acestei transformări poate fi ajustată în funcție de destinația cărnii.
Promovarea procesului de maturare poate duce la un proces de autoliză precoce; Această modificare a cărnii este cunoscută sub numele de fezandare. Macroul se practică de obicei în carnea tare, cu țesuturi dense, degresate, precum vânătoarea; După acest proces, carnea devine fragedă și are un gust deosebit de plăcut.
Clasificare. Clasificarea țesuturilor musculare se bazează pe două principii - morfofuncțional și histogenetic. În conformitate cu principiul morfofuncțional, în funcție de structura organelelor de contracție, țesuturile musculare sunt împărțite în două subgrupe.
Primul subgrup- tesut muscular striat (striat). (textus muscularis striatus).În citoplasma elementelor lor, filamente de miozină
Maturarea excesivă, chiar și atunci când carnea este depozitată în condiții absolut aseptice, poate duce la modificări organoleptice nedorite: culoarea se schimbă, carnea capătă un miros neplăcut, gustul devine acid amar. Această fază a maturării cărnii se numește autoliza cărnii și este un proces de descompunere a țesuturilor sub acțiunea enzimelor.
Masa termică se datorează proceselor chimice și biochimice care au loc în carne, însoțite de creșterea temperaturii cărnii; carnea fierbinte își schimbă culoarea și are gust dulce și acru. Lumina incandescentă este un proces de crăpare aleatoriu care are loc atunci când carnea fierbinte și umedă este depozitată fără a fi mai întâi răcită. Cu o ventilație puternică, efectele acestui proces pot fi reduse.
esti polimerizat permanent, formezi miofibrile existente permanent cu filamente de actina. Acestea din urmă sunt organizate în complexe caracteristice - sarcomere.În miofibrilele vecine, subunitățile structurale ale sarcomerelor sunt situate la același nivel și creează striatie transversala.
Al doilea subgrup- tesut muscular neted (nestriat). (textus muscularis nonstriatus). Aceste țesuturi se caracterizează prin faptul că, în afara contracției, filamentele de miozină sunt depolimerizate. În prezența ionilor de calciu, aceștia polimerizează și interacționează cu filamentele de actină. Miofibrilele formate in acest caz nu au striatii transversale: cu pete speciale, sunt reprezentate de fire uniform colorate (netede) pe toata lungimea.
Măcinarea cărnii are loc sub acțiunea bacteriilor aerobe și anaerobe care hidrolizează substanțele proteice. Ca urmare, carnea își schimbă aspectul, textura și mirosul, care devin specifice cărnii modificate. Începutul cărnii este marcat de apariția substanțelor volatile și de o schimbare a culorii care devine verde roșcat; În această fază se formează indol, skatol, baze azotate toxice etc.
Aspecte ale controlului sanitar veterinar în industria cărnii
Controlul sanitar veterinar se efectuează asupra animalelor vii pentru a detecta și exclude animalele bolnave și neucise. În plus, pentru a identifica eventualele boli infecțioase și parazitare, acestea sunt supuse controlului veterinar și a carcaselor și organelor interne ca urmare a sacrificării animalelor.
În conformitate cu principiul histogenetic, în funcție de sursele de dezvoltare (rudimente embrionare), țesuturile musculare și elementele musculare se împart în: somatice (miotomice), celomice (din placa mioepicardică a frunzei viscerale a splanhnotomului), mezenchimatoase (din rudimentul desmal ca parte a mezenchimului), neural (din tubul neural), epidermic (din ectodermul pielii și din placa precordală).
Carnea este cadrul striat, muscular al animalului, împreună cu toate țesuturile adezive, și anume țesutul conjunctiv, țesutul adipos, țesutul osos, vasele de sânge și nervii. Fondurile pentru locuințe sunt de obicei corpul unui animal sacrificat care a suferit o operațiune de curățare a suprafețelor, îndepărtarea organelor eviscerate cu comestibile și necomestibile și îndepărtarea membrelor.
De obicei, carcasele de bovine sunt împărțite în sferturi, porcii în jumătate, iar oile și păsările sunt lăsate intacte. După starea termică în care sunt livrate de către abatoare, carnea poate fi caldă, refrigerată, refrigerată și congelată.
9.2. ȚESUTURI MUSCULARE STRĂGATE
Există două varietăți principale de țesuturi striate (striate) - scheletice (miotomice) și cardiace (celomice).
9.2.1. Țesutul muscular scheletic
Histogenie. Sursa de dezvoltare a elementelor de țesut muscular scheletic (somatic). (textus muscularis striatus sceletalis) sunt celule stem ale miotomilor – promioblaste. Unii dintre ei se diferențiază în loc și participă la formarea așa-numiților mușchi autohtoni. Alte celule migrează de la miotomi la mezenchim. Sunt deja determinate, deși exterior nu diferă de alte celule mezenchimale. Diferențierea lor continuă în locurile de așezare a altor mușchi ai corpului. În timpul diferențierii, apar două linii celulare. Celulele uneia dintre linii se contopesc, formând simplaste alungite - tubuli musculari (miotuburi). În ele, diferențierea organelelor speciale - miofibrile (Fig. 9.1). În acest moment, în miotuburi se observă un reticul endoplasmatic granular bine dezvoltat. Miofibrilele sunt mai întâi localizate sub plasmalemă și apoi umplu cea mai mare parte a miotubului. Nucleii, dimpotrivă, din secțiunile centrale sunt deplasați spre periferie. Centrii celulari și microtubulii dispar complet. endo- granular
Carnea este produsă după ordinea individuală în conformitate cu scopul, tăind-o în zone anatomice. Tăierea rezultă în părțile anatomice ale celulei animale cu valori nutriționale diferite, care sunt împărțite în diferite calități și gamă.
Orez. 1 Transbordarea bovinelor. Reducerea se face pe baza următoarelor criterii: partea anatomică a carcasei sau categoria de carne, compoziția chimică a fiecărui tip de carne, inclusiv raportul dintre țesut muscular, os, grăsime, conjunctivă, valoarea nutritivă a cărnii. .
Pentru nevoi industriale, carnea este furnizată în carcase, jumătate de carcasă sau sferturi de carcasă. Carnea pentru catering poate fi clasificata in functie de tipul de preparat obtinut in timpul procesarii. Pentru piața internă, carnea de vită și mânzat adultă sunt clasificate în categorii de calitate, după cum urmează.
Orez. 9.1. Histogenia țesutului muscular scheletic (conform A. A. Klishov):
dar- promioblaste; b- miosimplast; în- tub muscular; G- mușchi matur
fibră. 1 - miosatellitocit; 2 - miezul miosimplastului; 3 - miofibrile
reticulul plasmatic este în mare măsură redus. Astfel de structuri definitive se numesc miosimplaste.
Pentru export, tăierea diferă în funcție de cerințele importatorului. Gradul de dezvoltare musculară și, în special, profilul pulpei și volumul mușchilor șosetelor și umerilor, dând mușchilor o clasă de cadru; gradul de dezvoltare a depozitelor de grăsime și, în special, a depozitelor de grăsime, precum și a grăsimilor din cavitatea abdominală și pelviană, care dau clasa de grăsime a carcasei. 
Orez. 2 Tăierea carcasei de porc.
muşchi; pisa; celuloză; mijloc cufăr; umerii; Gâtul pieptului; rulou; Încuietoare frontală; fiert înapoi; cap; picior din spate; în față. Carcasele de oi se clasifică în funcție de vârsta la care au fost sacrificate animalele: miel, îngrășat, oi și batalion. Deși este furnizat în ansamblu, componentele carcasei pot fi clasificate pentru piața internă în trei clase de calitate, în funcție de valoarea lor nutritivă.
Celulele dintr-o altă linie rămân independente și se diferențiază în miosatellitocite. Aceste celule sunt situate pe suprafața miosimplastelor. Miosatelitocitele, înmulțindu-se, fuzionează cu miosimplastele, participând astfel la crearea nucleară optimă.

Pentru export, conformația, culoarea cărnii, gradul de acoperire uniformă a grăsimii și randamentul la rasele timpurii specializate în producția de carne sunt estimate la 65%. Tăierea carcaselor de pasăre se face în diferite moduri, după piața de destinație. În general, carcasele sunt tăiate în următoarele părți anatomice: piept, spatulă, coloana vertebrală și spate sau piept, scripete și spate cu aripi.
Păsările sunt furnizate sub formă de carcase sau bucăți, refrigerate sau refrigerate. Este interzisă vânzarea păsărilor în stare dezghețată sau înghețată. În evaluarea calității cărnii, caracteristicile organoleptice joacă un rol important. Ele permit determinarea gradului de prospețime și identificarea anumitor defecte cauzate de starea animalelor sau de operațiunile de sacrificare, prelucrare și conservare. Principalele caracteristici organoleptice care determină gradul de prospețime al cărnii sunt: aspectul, culoarea, mirosul, textura, aspectul măduvei osoase și aspectul bulionului după fierbere.
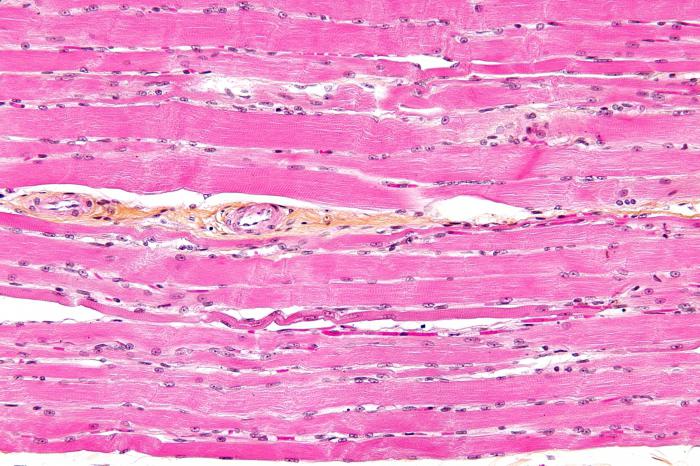
Orez. 9.2. Structura țesutului muscular striat (micrograf):
1 - fibre musculare; 2 - sarcolemă; 3 - sarcoplasmă și miofibrile; 4 - miezuri
miosimplast. Colorare - hematoxilină de fier
raportul sarcoplasmatic necesar sintezei proteinelor simplastice specifice.
Structura. Unitatea structurală principală a țesutului muscular scheletic este fibra musculară, care constă din miosimplast și miozatellocite acoperite de o membrană bazală comună (Fig. 9.2-9.4). Lungimea intregii fibre poate fi masurata in centimetri la o grosime de 50-100 microni. Complexul format din plasmolema miosimplast și membrana bazală se numește sarcolemă.
În plus, pentru a evalua prospețimea cărnii, se determină pH-ul acesteia și azotul ușor hidrolizabil. Reglementările sanitare veterinare prevăd absența agenților patogeni și a paraziților și paraziților alimentari, limitează numărul total de microbi mezofili din câmpul microscopic, numărul de bacterii care reduc sulfitul tisular, conținutul de pesticide și alți contaminanți.
După expunerea țesuturilor, se numește sarcom. Sarcoamele includ un grup de peste 40 de tipuri diferite de tumori. Sunt considerate relativ rare, reprezentând mai puțin de 1% din cancerele diagnosticate la adulți, cu aproximativ 000 de cazuri noi de sarcom de țesut moale și 400 de osteosarcoame în fiecare an. Sarcoamele țesuturilor moi pot apărea oriunde în corp, dar cele mai multe se formează în extremitățile inferioare, cel mai frecvent în coapse. Fibrosarcom conglomerat liposarcom osteosarcom angiosarcom miosarcom. . Sarcoamele apar, de asemenea, ca efecte secundare ale radioterapiei ca tratament pentru alte tipuri de cancer, dar sunt de obicei osteosarcoame.
Structura miosimplastului. Miosimplastul are mulți nuclei alungiți situati direct sub plasmalemă. Numărul lor într-un symplast poate ajunge la câteva zeci de mii (vezi Fig. 9.2). Organelele sunt situate la polii nucleelor sens general- Complexul Golgi și mici fragmente de agranulare reticulul endoplasmatic. Miofibrilele umplu partea principală a miosimplastului și sunt situate longitudinal (vezi Fig. 9.3).
Există anumite sugestii că există un risc crescut din cauza expunerii la anumite substanțe chimiceîn anumite industrii, dar studiile au arătat că acestea dau rezultate mixte și nu există o asociere clară. Un alt punct de vedere este că există o legătură între limfedemul cronic și o formă de sarcom numită limfangiosarcom. Factorii genetici care asociaza sau influenteaza dezvoltarea sarcomului sunt: sindromul Li-Fraumeni, neurofibromatoza, retinoblastomul ereditar. Deoarece cauzele exacte ale sarcomului sunt necunoscute, acesta nu poate fi prevenit.
Sarcomer- unitatea structurală a miofibrilei. Fiecare miofibrilă are discuri transversale întunecate și deschise cu refracție inegală (discuri A anizotrope și discuri I izotrope). Fiecare miofibrilă este înconjurată de bucle dispuse longitudinal și anastomozatoare ale reticulului endoplasmatic agranular - reticulul sarcoplasmatic. Sarcomerele învecinate au o structură de frontieră comună - linia Z (Fig. 9.5). Este construit sub forma unei rețele de molecule fibrilare proteice, printre care alfa-actinina joacă un rol semnificativ. Capetele filamentelor de actină sunt conectate la această rețea. Din liniile Z vecine, filamentele noi de actină sunt direcționate spre centrul sarcomerului, dar nu ajung la mijlocul acestuia. Filamentele de actină sunt conectate la linia Z și filamentele de miozină
Cu toate acestea, este recomandat să evitați substanțele chimice enumerate mai sus. Din păcate, nu există un test de screening pentru sarcoamele țesuturilor moi. Programele de screening sunt concepute pentru a detecta precoce bolile comune sau fatale. Avand in vedere ca sarcoamele tesuturilor moi sunt foarte rare, ar fi dificil sa se realizeze un program de screening pentru intreaga populatie. De asemenea, este foarte dificil să se dezvolte un singur program de screening care să acopere toate tipurile de sarcom de țesut moale.
Simptomele sarcomului de țesut moale depind de zona tumorii. Deoarece majoritatea se formează la nivelul extremităților, acești pacienți se vor plânge de o masă sau nodul la nivelul extremităților, cu sau fără denivelări. În funcție de locație și dimensiune, aceasta poate cauza sau nu durere. Dacă tumora se formează în cavitatea abdominală, aceasta va atinge o dimensiune semnificativă până când este detectată sau provoacă durere. Tumorile care apar în tractul gastrointestinal pot provoca diaree, sânge în scaun sau dureri abdominale.

Orez. 9.3. Schema structurii ultramicroscopice a miosimplastului (după R. V. Krstic, cu modificări) (a): 1 - sarcomer; 2 - disc anizotrop (banda A); 2a - disc izotrop (banda I); 3 - linia M (mezofragmă) în mijlocul discului anizotrop; 4 - linia Z (telofragmă) în mijlocul unui disc izotrop; 5 - mitocondrii; 6 - reticulul sarcoplasmatic; 6a - rezervor final; 7 - tub transversal (T-tube); 8 - triadă; 9 - sarcolemă; b- schema dispozitiei spatiale a mitocondriilor din simplast. Planurile superioare și inferioare ale figurii limitează măsura sarcomului de disc izotrop (conform L. E. Bakeeva, V. P. Skulachev, Yu. S. Chentsov); în- endomisiu. Micrografie electronică cu scanare, mărire 2600 (pregătirea lui Yu. A. Khoroshkov): 1 - fibre musculare; 2 - fibrile de colagen
molecule de nebuline fibrilare inextensibile. În mijlocul discului întunecat al sarcomerului se află o rețea construită din miomiozină. Formează o linie M în secțiune transversală. Capetele filamentelor de miozină sunt fixate la nodurile acestei linii M. Celelalte capete ale lor sunt îndreptate spre liniile Z și

Orez. 9.4. Zona superficială a miosimplastului și miosatelitecitului. Micrografie electronică, mărire 10.000 (preparat de V. L. Goryachkina și S. L. Kuznetsov): 1 - membrana bazală; 2 - plasmalema; 3 - miezul miosimplastului; 4 - nucleul mios-telitocitului; 5 - miofibrile; 6 - tubuli ai reticulului endoplasmatic (sarcoplasmatic) agranular; 7 - mitocondrii; 8 - glicogen

Orez. 9.5. Sarcomer (schemă):
1 - linia Z; 2 - linia M; 3 - filamente de actină; 4 - filamente de miozină; 5 - molecule de titină fibrilare (după B. Alberts, D. Bray, J. Lewis și alții, cu modificări)

Orez. 9.6. Modificări conformaționale care implică deplasarea reciprocă a filamentelor de actină și miozină:
a-c- modificări succesive ale relaţiilor spaţiale. 1 - actină; 2 - capul moleculei de miozină (după B. Alberts, D. Bray, J. Lewis et al., cu modificări)
sunt situate între filamentele de actină, dar nu ajung nici la liniile Z în sine. În același timp, aceste capete sunt fixate în raport cu liniile Z prin molecule de proteine gigantice extensibile ale titinei.
Moleculele de miozină au o coadă lungă și două capete la unul dintre capete. Odată cu creșterea concentrației de ioni de calciu în zona de atașare a capetelor (secțiunea balamalei), molecula își schimbă configurația (Fig. 9.6). În acest caz (deoarece filamentele de actină sunt situate între filamentele de miozină), capetele de miozină se leagă de actină (cu participarea proteinelor auxiliare - tropomiozina și troponina). Apoi capul miozinei se înclină și trage molecula de actină spre linia M. Liniile Z converg, sarcomerul se scurtează.
Rețelele de alfa-actinină ale liniilor Z ale miofibrilelor învecinate sunt conectate între ele prin filamente intermediare. Ele se apropie de suprafața interioară a plasmolemei și sunt fixate în stratul său cortical, astfel încât sarcomerele tuturor miofibrilelor să fie situate la același nivel. Acest lucru creează, atunci când este observat la microscop, impresia unei striații transversale a întregii fibre.
Cisternele reticulului endoplasmatic agranular servesc ca sursă de ioni de calciu. Ele sunt extinse de-a lungul miofibrilelor în apropierea fiecărui sarcomer și formează un reticul sarcoplasmatic. În ea se acumulează ionii de calciu atunci când miosimplastul este într-o stare relaxată. La nivelul liniilor Z (la amfibieni) sau la granița discurilor A și I (la mamifere), tubii rețelei își schimbă direcția și sunt localizați transversal, formând cisterne terminale sau laterale (L) extinse.
De la suprafața până la adâncimea miosimplastului, plasmolema formează tuburi lungi care se deplasează transversal (tubuli T) la nivelul limitelor dintre discurile întunecate și cele deschise. Când miosimplastul primește un semnal de inițiere a contracției, se deplasează de-a lungul plasmalemei sub forma unui potențial de acțiune și se propagă la membrana tubului T. Deoarece această membrană este aproape de membranele reticulului sarcoplasmatic, starea acestuia din urmă se schimbă, calciul este eliberat din cisternele rețelei și interacționează cu complexele actină-miozină (se contractă). Când potențialul de acțiune dispare, calciul se acumulează din nou în tubii rețelei și contracția miofibrilelor se oprește. Energia este necesară pentru a dezvolta forța de contracție. Este eliberat prin conversia ATP în ADP. Rolul ATPazei este îndeplinit de miozină. Sursa de ATP este în principal mitocondriile, deci sunt situate direct între miofibrile.
Incluziunile de mioglobină și glicogen joacă un rol important în activitatea miosimplastelor. Glicogenul servește ca sursă de energie necesară nu numai pentru a efectua munca musculară, ci și pentru a menține echilibrul termic al întregului organism. Mioglobina leagă oxigenul atunci când mușchiul este relaxat și sângele curge liber prin vasele mici de sânge. În timpul contracției musculare, vasele sunt comprimate, iar oxigenul stocat este eliberat și participă la reacții biochimice.
Miosatellocite. Aceste celule nediferențiate sunt sursa regenerării țesutului muscular. Ele sunt adiacente suprafeței miosimplastului, astfel încât membranele lor plasmatice sunt în contact (vezi Fig. 9.1, 9.4). Miosatelitocitele sunt mononucleare, nucleii lor întunecați sunt ovali și mai mici decât la simplaste. Au toate organelele de importanță generală (inclusiv centrul celular).
Tipuri de fibre musculare. Diferiți mușchi (precum organele) funcționează în diferite condiții biomecanice. Prin urmare, fibrele musculare din compoziția diferiților mușchi au forță, viteză și durata de contracție diferite, precum și oboseală. Activitatea enzimelor din ele este diferită și sunt prezentate în diferite forme izomerice. Diferite în ele și conținutul de enzime respiratorii - glicolitice și oxidative.

Orez. 9.7. Activitatea succinat dehidrogenazei în fibrele musculare de diferite tipuri (preparatul VF Chetvergov, tratament conform Nahlas și colab.): 1 - mare; 2 - scăzut; 3 - mediu
După raportul dintre miofibrile, mitocondrii și mioglobină, se disting fibrele albe, roșii și intermediare. În funcție de caracteristicile funcționale, fibrele musculare sunt împărțite în rapide, lente și intermediare, ceea ce este determinat de organizarea moleculară a miozinei. Printre izoformele sale, există două principale - „rapid” și „lent”. La stabilirea reacțiilor histochimice, acestea sunt identificate prin activitatea ATP-azei. Aceste proprietăți se corelează cu activitatea enzimelor respiratorii. Procesele glicolitice predomină de obicei în fibrele rapide, sunt bogate în glicogen, au mai puțină mioglobină, de aceea se numesc albe. În fibrele lente, dimpotrivă, activitatea enzimelor oxidative este mai mare, sunt mai bogate în mioglobină și arată mai roșu.
Alături de alb și roșu, există și fibre intermediare. În compoziția majorității mușchilor scheletici, fibrele de diferite tipuri histochimice sunt aranjate într-un model mozaic (Fig. 9.7).
Proprietățile fibrelor musculare se modifică odată cu modificările sarcinilor - sportive, profesionale și, de asemenea, în condiții extreme (imponderabilitate). La revenirea la activitățile normale, astfel de modificări sunt reversibile. În unele boli (atrofie musculară, distrofie, consecințe ale denervației), fibrele musculare cu proprietăți inițiale diferite se schimbă diferit. Acest lucru vă permite să clarificați diagnosticul, pentru care sunt examinate probe de biopsie ale mușchilor scheletici.
Regenerare. Nucleii miosimplastelor nu se pot diviza, deoarece nu există centri celulari în sarcoplasmă. Elementele cambiale sunt miozatellocite. Pe măsură ce organismul crește, se divid, iar celulele fiice se contopesc cu miosimplastele. La sfârșitul creșterii, reproducerea miosatellitocitelor se estompează. După deteriorarea fibrei musculare la o anumită distanță de locul leziunii, aceasta se prăbușește și se fragmentează
esti fagocitat de macrofage. Repararea țesuturilor se realizează prin două mecanisme: hipertrofia compensatorie a symplast în sine și proliferarea miosatellitocitelor. În simplast sunt activate reticulul endoplasmatic granular și complexul Golgi. Există o sinteză de substanțe necesare refacerii sarcoplasmei și miofibrilelor, precum și asamblarea membranelor, astfel încât integritatea membranei plasmatice să fie restabilită. Capătul deteriorat al miosimplastului se îngroașă, formând un rinichi muscular. Miosatelitocitele, conservate în apropierea leziunii, se divid. Unii dintre ei migrează în rinichiul muscular și se integrează în acesta, alții fuzionează (la fel ca mioblastele în timpul histogenezei) și formează noi miotuburi care se dezvoltă în fibre musculare.
9.2.2. Mușchi scheletic ca organ
Transmiterea forțelor de contracție către schelet se realizează prin intermediul tendoanelor sau atașării musculare direct la periost. La capătul fiecărei fibre musculare, plasmalema formează proeminențe înguste adânci. Fibrele subțiri de colagen pătrund în ele din partea tendonului sau a periostului. Acestea din urmă sunt împletite spiralat cu fibre reticulare. Capetele fibrelor merg la membrana bazală, intră în ea, se întorc și, la ieșire, împletesc din nou fibrele de colagen țesut conjunctiv.
Între fibrele musculare sunt straturi subțiri de țesut conjunctiv fibros lax - endomisiu.În ea sunt țesute fibrele de colagen ale foii exterioare a membranei bazale (vezi Fig. 9.3, c), ceea ce contribuie la unificarea eforturilor în timpul contracției miosimplastelor. Straturi mai groase de țesut conjunctiv lax înconjoară mai multe fibre musculare, formându-se perimisiuşi împărţirea muşchiului în mănunchiuri. Mai multe fascicule sunt combinate în grupuri mai mari, separate prin straturi mai groase de țesut conjunctiv. Țesutul conjunctiv care înconjoară suprafața unui mușchi se numește epimisio.
Vascularizarea. Arterele intră în mușchi și se răspândesc prin straturile de țesut conjunctiv, devenind treptat mai subțiri. Ramurile de ordinul al cincilea sau al șaselea formează arteriole în perimisium. Capilarele sunt localizate în endomisiu. Ele merg de-a lungul fibrelor musculare, anastomozându-se între ele. Venulele, venele și vasele limfatice trec pe lângă vasele aferente. Ca de obicei, lângă vase există multe mastocite implicate în reglarea permeabilității peretelui vascular.
Inervație.În mușchi s-au găsit fibre eferente (motorie), aferente (senzoriale), precum și fibre nervoase autonome nemielinice. ramură celula nervoasa, aducând impulsul de la motoneuron măduva spinării, ramuri în perimisium. Fiecare dintre ramurile sale pătrunde în membrana bazală și formează terminale pe plasmolema la suprafața symplast, participând la organizarea așa-numitei plăci motorii (vezi Capitolul 10, Fig. 10.18). La admitere

Orez. 9.8. Un fragment dintr-un fus muscular care conține fibre musculare cu lanț nuclear (a) și cu pungă nucleară (b) (schemă după G. S. Katinas): 1 - nuclei; 2 - miofibrile (nu sunt prezentate organele de importanță generală)
al impulsului nervos, de la terminale este eliberată acetilcolina, un mediator, ceea ce provoacă excitație (potențial de acțiune), care se propagă de aici de-a lungul plasmolemei miosimplast.
Deci, fiecare fibră musculară este inervată independent și înconjurată de o rețea de hemocapilare, formând un complex numit myon.
Se numește un grup de fibre musculare inervate de un singur neuron motor unitate neuromusculară. Fibrele musculare care aparțin unei unități neuromusculare nu se află una lângă alta, ci sunt situate într-un mozaic printre fibrele aparținând altor unități.
Terminațiile nervoase senzoriale nu sunt localizate pe fibrele musculare de lucru (extrafusale), ci sunt asociate cu fibre musculare specializate din așa-numitele lanțuri musculare.
tenah (cu fibre musculare intrafusale), care sunt situate în perimisium.
fibre musculare intrafusale. Fibrele musculare intrafusale ale fusurilor sunt mult mai subțiri decât lucrătorii. Există două tipuri de ele - fibre cu o pungă nucleară și fibre cu un lanț nuclear (Fig. 9.8). Nucleii din ambele sunt rotunjite și situate în grosimea simplastului și nu aproape de suprafața acestuia. În fibrele cu o pungă nucleară, nucleii simplast formează grupuri în partea mijlocie îngroșată. În fibrele cu lanț nuclear, nu se formează nicio îngroșare în partea mijlocie a simplastului; nucleii se află aici longitudinal unul după altul. Organelele de importanță generală sunt situate lângă grupurile de nuclee.
Miofibrilele sunt situate la capetele simplastelor. Sarcolema fibrelor se conectează la capsula fusului neuromuscular, care este compusă din țesut conjunctiv fibros dens. Fiecare fibră musculară a fusului este înfășurată în spirală în jurul terminalului unei fibre nervoase sensibile. Ca urmare a contracției sau relaxării fibrelor musculare care lucrează, tensiunea capsulei de țesut conjunctiv a fusului se modifică, iar tonusul fibrelor musculare intrafusale se modifică în consecință. Ca rezultat, terminațiile nervoase sensibile sunt excitate, înfășurându-le în jurul lor, iar impulsurile nervoase aferente apar în regiunea terminalelor. Fiecare miosimplast are și propria sa placă motorie. De aceea fibrele musculare intrafuzale sunt în permanență în tensiune, ajustându-se la lungimea abdomenului muscular în ansamblu.
9.2.3. țesut muscular cardiac
Histogenie și tipuri de celule. Surse de dezvoltare a țesutului muscular striat cardiac (textus muscular striatus cardiacus)- secțiuni simetrice ale frunzei viscerale a splanhnotomului în partea cervicală a embrionului - plăci mioepicardice. Dintre acestea, se diferențiază și celulele mezoteliului epicardului. Celulele originale ale țesutului muscular cardiac - cardiomioblaste- caracterizat printr-o serie de caracteristici: celulele sunt turtite, contin un nucleu mare, citoplasma usoara, saraca in ribozomi si mitocondrii. În viitor, are loc dezvoltarea complexului Golgi, un reticul endoplasmatic granular. Structurile fibrilare se găsesc în cardiomioblaste, dar nu miofibrile. Celulele au un potențial proliferativ ridicat.
După o serie de cicluri mitotice, cardiomioblastele se diferențiază în cardiomiocite,în care începe sarcomerogeneza (fig. 9.9). În citoplasma cardiomiocitelor, numărul de polizomi, tubuli ai reticulului endoplasmatic granular crește, se acumulează granule de glicogen, iar volumul complexului de actomiozină crește. Cardiomiocitele sunt reduse, dar nu își pierd capacitatea de proliferare și diferențiere în continuare. Dezvoltarea aparatului contractil în perioadele embrionare târzii și postnatale are loc prin adăugarea de noi sarcomere și stratificarea miofilamentelor nou sintetizate.
Diferențierea cardiomiocitelor este însoțită de o creștere a numărului de mitocondrii, distribuția acestora la polii nucleelor și între miofibrile și decurge în paralel cu specializarea suprafețelor celulare în contact. Cardiomiocitele prin contacte „cap la cap”, „cap la cap” formează fibre musculare cardiace și, în general, țesutul este o structură de rețea. Unele cardiomiocite aflate în stadiile incipiente ale cardiomiogenezei sunt contractilo-secretorii. Ulterior, ca urmare a diferențierii divergente, apar miocite „întunecate” (contractile) și „luminoase” (conductoare), în care granulele secretoare dispar, în timp ce rămân în miocitele atriale. Așa se formează diferența de cardiomiocite endocrine. Aceste celule conțin un nucleu situat central cu cromatina dispersată, unul sau doi nucleoli. În citoplasmă, un reticul endoplasmatic granular, dictiozomi ai complexului Golgi, sunt bine dezvoltati, în strânsă legătură cu elementele cărora există numeroase granule secretoare cu un diametru de aproximativ 2 microni, care conțin material electrodens. În viitor, granulele secretoare se găsesc sub sarcolemă și sunt eliberate în spațiul extracelular prin exocitoză.
În general, în timpul histogenezei, apar cinci tipuri de cardiomiocite - de lucru (contractile), sinusale (stimulator cardiac), tranzitorii, conductoare și, de asemenea, secretorii. Cardiomiocite de lucru (contractile). formează propriile lanțuri (Fig. 9.10). Ei sunt cei care, scurtând, asigură forța de contracție a întregului mușchi al inimii. Cardiomiocitele de lucru sunt capabile

Orez. 9.9. Histogenia țesutului muscular cardiac (schema conform lui P. P. Rumyantsev): dar- cardiomiocite în peretele tubului cardiac; b - cardiomiocite în embriogeneză târzie; în- cardiomiocite în perioada postnatală. 1 - cardiomiocit; 2 - cardiomiocitul care se divide mitotic; 3 - miofilamente și miofibrile
transmite semnale de control unul altuia. Cardiomiocite sinusale (stimulator cardiac). sunt capabili să schimbe automat starea de contracție în starea de relaxare într-un anumit ritm. Celulele percep semnale de control de la fibrele nervoase, ca răspuns la care schimbă ritmul activității contractile. Cardiomiocitele sinusale (pacemaker) transmit semnale de control cardiomiocite de tranziție, iar acesta din urmă - la cardiomiocite conducătoare și de lucru. Cardiomiocite conducătoare formează lanțuri de celule conectate la capetele lor și sunt situate sub endo-

Orez. 9.10. Structura țesutului muscular cardiac (micrograf). Colorare - hematoxilină de fier:
1 - nucleul unui cardiomiocit; 2 - un lanț de cardiomiocite; 3 - introduceți discuri
card. Prima celulă din lanț primește semnale de control de la cardiomiocitele sinusurilor și le transmite altor cardiomiocite conducătoare. Celulele care completează lanțul transmit un semnal către lucrători prin cardiomiocite de tranziție. Cardiomiocitele secretoareîndeplini o funcție specială. Ele produc hormonul peptidic cardiodilatina, care circulă în sânge sub formă de cardionatrino, provoacă contracția miocitelor netede ale arteriolelor, crește fluxul sanguin renal, accelerează filtrarea glomerulară și excreția de sodiu. Toate cardiomiocitele sunt acoperite cu o membrană bazală.
Structura cardiomiocitelor contractile (de lucru). Celulele au o formă alungită (100-150 microni), apropiată de cilindrică. Capetele lor sunt legate între ele, astfel încât lanțurile de celule alcătuiesc așa-numitele fibre funcționale (până la 20 de microni grosime). În zona contactelor celulare, se formează așa-numitele discuri intercalate (Fig. 9.10). Cardiomiocitele se pot ramifica și forma o rețea spațială. Suprafețele lor sunt acoperite cu o membrană bazală, în care sunt țesute fibre reticulare și de colagen din exterior. Nucleul unui cardiomiocite (uneori sunt doi) este oval și se află în partea centrală a celulei. Câteva organele de importanță generală sunt concentrate la polii nucleului, cu excepția reticulului endoplasmatic agranular și a mitocondriilor.
Organele speciale care asigură contracția se numesc miofibrile. Sunt ușor separați unul de celălalt, se pot despărți. Structura lor este similară cu cea a miofibrilelor miosimplastului fibrei musculare scheletice. Fiecare mitocondrie este localizată în întregul sarcomer. De la suprafața plasmolemei, tubulii T sunt direcționați adânc în cardiomiocit, situat la nivelul liniei Z. Membranele lor sunt apropiate
contactul cu membranele reticulului endoplasmatic neted (sarcoplasmatic). Ansele acestora din urmă sunt extinse de-a lungul suprafeței miofibrilelor și au prelungiri laterale (sisteme L), care împreună cu tubii T formează triade sau diade (Fig. 9.11, a). În citoplasmă există incluziuni de glicogen și lipide, în special multe incluziuni de mioglobină. Mecanismul de contracție al cardiomiocitelor este același cu cel al miosimplastului.
Organizarea cardiomiocitelor în țesut. Cardiomiocitele se conectează între ele într-un mod de la capăt la capăt. Aici se formează discuri intercalate: aceste zone arată ca niște plăci subțiri la mărire medie microscop luminos. De fapt, capetele cardiomiocitelor au o suprafață neuniformă, astfel încât proeminențele unei celule intră în depresiunile celeilalte. Secțiunile transversale ale proeminențelor celulelor învecinate sunt legate între ele prin interdigitări și desmozomi (Fig. 9.11, b).

Orez. 9.11. Structura unui cardiomiocit: dar- schema (conform lui Yu. I. Afanasyev și V. L. Goryachkina); b- micrografie electronică a discului de inserție. Mărire 20 000. 1 - miofibrile; 2 - mitocondrii; 3 - retea sarcotubulara; 4 - tubuli T; 5 - membrana bazala; 6 - lizozom; 7 - introduceți disc; 8 - desmozom; 9 - zona de atașare a miofibrilelor; 10 - contacte întrerupte; 11 - glicogen
O miofibrilă se apropie de fiecare desmozom din partea laterală a citoplasmei, iar capătul acesteia este fixat în complexul desmo-plakin. Astfel, în timpul contracției, forța unui cardiomiocite este transferată către alta. Suprafetele laterale proeminențele cardiomiocitelor sunt unite prin nexusuri (joncțiuni gap). Acest lucru creează conexiuni metabolice între ele și asigură sincronia contracțiilor.
Regenerare.În histogeneza țesutului muscular cardiac, cambiul nu apare. Prin urmare, regenerarea tisulară are loc pe baza proceselor hiperplazice intracelulare. În același timp, pentru cardiomiocitele de mamifere, primate și oameni, procesul de poliploidie este caracteristic.
zarea. De exemplu, la maimuțe, nucleele de până la 50% din cardiomiocitele diferențiate terminal devin tetra- și octoploide. Cardiomiocitele poliploide apar din cauza mitozei, ceea ce duce la multinucleare. În condiții de patologie a sistemului cardiovascular uman (reumatism, malformații cardiace congenitale, infarct miocardic etc.), regenerarea intracelulară, poliploidizarea nucleelor și apariția cardiomiocitelor multinucleare joacă un rol important în compensarea deteriorarii cardiomiocitelor.
9.3. ȚESUT MUSCULAR NETED
Există trei grupuri de țesuturi musculare netede (nestriate). (textus muscularis nonstriatus)și celule: celule mezenchimale, neuronale și mioepiteliale.
9.3.1. Muşchi origine mezenchimală
Histogenie. Acest țesut este împărțit în două tipuri: visceral și vascular. În histogeneza embrionară, este chiar dificil să se distingă precursorii mezenchimale ai fibroblastelor de miocitele netede chiar și prin microscopie electronică. În miocitele netede slab diferențiate, se dezvoltă un reticul endoplasmatic granular, complexul Golgi. Filamentele subțiri sunt orientate de-a lungul axei lungi a celulei. Pe măsură ce celula se dezvoltă, mărimea celulei și numărul de filamente din citoplasmă cresc. Treptat, volumul citoplasmei ocupate de filamentele contractile crește, localizarea lor în citoplasmă devine din ce în ce mai ordonată. Activitatea proliferativă a miocitelor netede scade treptat în timpul miogenezei. Aceasta apare ca urmare a creșterii duratei ciclului celular, a ieșirii celulelor din ciclul de reproducere și a trecerii la o stare diferențiată. Diferențiând, sintetizează componentele matricei extracelulare, colagenul membranei bazale, precum și elastina. În celulele definitive (miocite), capacitatea de sinteză este redusă, dar nu dispare complet.
Structura și funcția celulelor. Un miocit neted este o celulă fuziformă de 20–500 µm lungime și 5–8 µm lățime. Nucleul este în formă de tijă, situat în partea centrală. Când miocitul se contractă, nucleul său se îndoaie și chiar se răsucește (Fig. 9.12-9.14).
Structura miocitelor netede definitive (leiomiocite), care fac parte din organe interneși pereții vaselor, are multe în comun, dar în același timp se caracterizează prin heteromorfie. Deci, în pereții venelor și arterelor, se găsesc miocite ovoide, fusiforme, procesate de 10-40 microni lungime, ajungând uneori până la 140 microni. Cel mai lung mio-
citele ajung în peretele uterului - până la 500 de microni. Diametrul miocitelor variază de la 2 la 20 de microni. În funcție de natura proceselor de biosinteză intracelulară, se disting miocitele contractile și secretoare. Primele sunt specializate în funcția de contracție, dar păstrează în același timp și activitate secretorie.
Miocitele secretoare seamănă cu fibroblastele în ultrastructură, dar conțin în citoplasmă mănunchiuri de miofilamente subțiri situate la periferia celulei. În citoplasmă, complexul Golgi, reticulul endoplasmatic granular, multe mitocondrii, granule de glicogen, ribozomi liberi și polizomi sunt bine dezvoltate. În funcție de gradul de maturitate, astfel de celule sunt clasificate ca nediferențiate. Filamentele de actină formează o rețea tridimensională în citoplasmă, alungită în principal longitudinal. Capetele filamentelor sunt atașate unele de altele și de plasmalemă prin proteine speciale de reticulare. Aceste zone sunt clar vizibile pe micrografiile electronice ca corpuri dense. Monomerii miozinei sunt localizați lângă filamentele de actină. Plasmalema formează invaginări - caveole, în care sunt concentrați ionii de calciu. Semnalul de contractare vine de obicei prin fibrele nervoase. Mediatorul care este eliberat de la terminalele lor modifică permeabilitatea membranei plasmatice. Are loc eliberarea ionilor de calciu, ceea ce presupune atât polimerizarea miozinei, cât și interacțiunea miozinei cu actina.
Există o retragere a miofilamentelor de actină între miozi-

Orez. 9.12. Structura unui miocit neted (schemă):
dar, în- la relaxare; b, d- la cea mai mare reducere; G- cu contractie incompleta; v-d- imagini mărite ale zonelor încercuite de cadre pe fragmente darși b. 1 - plasmalema; 2 - corpuri dense; 3 - miez; 4 - endoplasma; 5 - complexe contractile; 6 - mitocondrii; 7 - membrana bazala; 8 - mio-filamente (subțiri) de actină; 9 - miofilamente (groase) de miozină

Orez. 9.13. Ultrastructura unui miocit neted diferențiat în peretele bronhiei:
1 - miez; 2 - citoplasmă cu miofilamente; 3 - Complexul Golgi, mărire 35.000 (preparat de A. L. Zashikhin)
pete noi, dense, se apropie unele de altele, forța este transferată plasmalemei și întreaga celulă se scurtează (vezi Fig. 9.12). Când semnalele de la sistemul nervos se opresc, ionii de calciu se deplasează din citoplasmă în caveole și în tubii reticulului endoplasmatic, miozina se depolimerizează și „miofibrilele” se dezintegrează. Contracția se oprește. Astfel, complexele de actinomiozină există în miocitele netede numai în timpul contracției în prezența ionilor liberi de calciu în citoplasmă.
Miocitele sunt înconjurate de o membrană bazală. În unele zone, în el se formează „ferestre”, astfel încât membranele plasmatice ale miocitelor vecine se apropie una de cealaltă. Aici se formează legături și între celule apar nu numai conexiuni mecanice, ci și metabolice. Fibrele elastice și reticulare trec peste „acoperirile” din membrana bazală dintre miocite, unind celulele într-un singur complex tisular. Miocitele netede sintetizează proteoglicani, glicoproteine, procolagen, proelastină, din care se formează colagen și fibre elastice și o componentă amorfă a matricei extracelulare. Interacțiunea miocitelor se realizează cu ajutorul punților citoplasmatice, invaginărilor reciproce, lexurilor, desmozomilor, zonelor de contact membranar ale suprafețelor miocitelor.
Regenerare.Țesutul muscular neted de tip visceral și vascular este foarte sensibil la factorii extremi. În miocitele activate, nivelul proceselor de biosinteză crește, a căror expresie morfologică este sinteza proteinelor contractile, mărirea și hipercromatoza nucleului, hipertrofia nucleolului, creșterea raportului nuclear-citoplasmatic, creșterea numărului de ribozomi și polizomi liberi, activare

Orez. 9.14. Structura țesutului muscular neted (schemă volumetrică) (conform R. V. Krstic, cu modificări):
1 - miocite netede fuziforme; 2 - citoplasma miocitelor; 3 - nuclee de miocite; 4 - plasmalema; 5 - membrana bazala; 6 - vezicule pinocitare superficiale; 7 - conexiuni intercelulare; 8 - terminație nervoasă; 9 - fibrile de colagen; 10 - microfilamente
enzime, fosforilare aerobă și anaerobă, transport membranar. Regenerarea celulară se realizează atât datorită celulelor diferențiate care au capacitatea de a intra în ciclul mitotic, cât și datorită activării elementelor cambiale (miocite de volum mic). Sub acțiunea unui număr de factori dăunători, se observă o transformare fenotipică a miocitelor contractile în cele secretoare. Această transformare este adesea observată cu deteriorarea intimei vaselor, formarea hiperplaziei intimale în timpul dezvoltării aterosclerozei.

Orez. 9.15. Ultrastructura miopigmentocitelor (preparat de N. N. Sarbaeva): 1 - miez; 2 - miofilamente, mărire 6000
9.3.2. Țesut muscular de tip mezenchimatos în compoziția organelor
Miocitele sunt combinate în mănunchiuri, între care există straturi subțiri de țesut conjunctiv. Fibrele reticulare și elastice care înconjoară miocitele sunt țesute în aceste straturi. În straturi se află vase de sânge și fibre nervoase. Terminalele acestora din urmă nu se termină direct pe miocite, ci între ele. Prin urmare, după sosirea unui impuls nervos, mediatorul se răspândește difuz, excitând multe celule simultan. Țesutul muscular neted de origine mezenchimală este reprezentat în principal în pereții vaselor de sânge și în multe organe interne goale.
Țesutul muscular neted din compoziția unor organe specifice are proprietăți funcționale diferite. Acest lucru se datorează faptului că pe suprafața organelor există diferiți receptori pentru anumite substanțe biologic active. Prin urmare, răspunsul lor la multe medicamente nu este același. Este posibil ca diferite proprietăți funcționale ale țesuturilor să fie, de asemenea, asociate cu organizarea moleculară specifică a filamentelor de actină.
9.3.3. Țesut muscular de origine neuronală
Țesutul muscular al irisului și al corpului ciliar aparține celui de-al patrulea tip de țesuturi contractile. Miocitele acestui țesut se dezvoltă din celulele rudimentului neural ca parte a peretelui interior al ocularului. Consecutiv

Orez. 9.16. Celulele mioepiteliale din secțiunea terminală a glandei salivare (schemă după G. S. Katinas):
dar- secțiune transversală; b- vedere de la suprafata. 1 - nuclee de mioepiteliocite; 2 - procesele mioepiteliocitelor; 3 - nucleele epiteliocitelor secretoare; 4 - membrană bazală
La vertebrate, elementele musculare ale irisului prezintă o varietate de diferențieri divergente. Astfel, tesutul mioneural la reptile si pasari este reprezentat de fibre multinucleare striate, care sunt foarte asemanatoare cu muschii de tip scheletic. La mamifere și oameni, principala unitate structurală și funcțională a mușchilor irisului este un miocit mononuclear neted sau miopigmentocit. Acestea din urmă au un corp pigmentat care conține un nucleu, care se desfășoară dincolo de limitele părții contractile fuziforme (Fig. 9.15).
Citoplasma celulelor contine număr mare mitocondriile și granulele pigmentare, care sunt similare ca mărime și formă cu granulele epiteliale pigmentare. Miofilamentele din miopigmentocite sunt împărțite în subțiri (7 nm) și groase (1,5 nm), ca dimensiune și locație seamănă cu miofilamentele miocitelor netede. Fiecare miopigmentocit este înconjurat de o membrană bazală. În apropierea proceselor citoplasmatice ale miocitelor se găsesc fibre nervoase nemielinice. În funcție de direcția proceselor (perpendiculară sau paralelă cu marginea pupilei), miocitele formează doi mușchi - constrângând și extinzând pupila.
Regenerare. Puține lucrări arată o activitate regenerativă scăzută după leziune sau absența acesteia.
9.3.4. Celule musculare de origine epidermică
Celulele mioepiteliale se dezvoltă din mugurele epidermic. Se găsesc în glandele sudoripare, mamare, salivare și lacrimale și au precursori comuni cu celulele lor secretoare. Mioepitelial
celulele sunt direct adiacente celulelor epiteliale propriu-zise și au o membrană bazală în comun cu acestea. În timpul regenerării, acele și alte celule sunt, de asemenea, restaurate din precursori comuni nediferențiați. Majoritatea celulelor mioepiteliale au formă stelată. Aceste celule sunt adesea numite celule coș: procesele lor acoperă secțiunile terminale și canalele mici ale glandelor (Fig. 9.16). Nucleul și organitele de importanță generală sunt localizate în corpul celular, iar aparatul contractil este situat în procese, organizate ca și în celulele țesutului muscular de tip mezenchimal.
întrebări de testare
1. Clasificarea genetică a țesuturilor musculare. Unități structurale și funcționale ale diferitelor tipuri de țesut muscular.
2. Țesutul muscular scheletic striat: dezvoltarea, structura, baza morfologică a contracției musculare. Regenerare.
3. Țesut muscular cardiac striat: dezvoltare, specificul structural al diferitelor tipuri de cardiomiocite, regenerare.
4. Soiuri de miocite netede: surse de dezvoltare, topografie în organism, regenerare.
Histologie, embriologie, citologie: manual / Yu. I. Afanasiev, N. A. Yurina, E. F. Kotovsky și alții - ed. a 6-a, revizuită. si suplimentare - 2012. - 800 p. : bolnav.
Ele îndeplinesc o funcție foarte importantă în organismele ființelor vii - formează și căptușesc toate organele și sistemele lor. De o importanță deosebită printre aceștia este tocmai mușchiul, deoarece importanța sa în formarea cavităților exterioare și interioare ale tuturor părților structurale ale corpului este o prioritate. În acest articol, vom lua în considerare ce este țesutul muscular neted, caracteristicile sale structurale, proprietățile.
Varietăți ale acestor țesături
Există mai multe tipuri de mușchi în compoziția corpului animalului:
- striat;
- țesut muscular neted.
Ambele au propriile trăsături caracteristice ale structurii, funcțiilor îndeplinite și proprietăților expuse. În plus, sunt ușor de distins unul de celălalt. La urma urmei, ambele au propriul lor model unic, care se formează datorită componentelor proteice care alcătuiesc celulele.
Cu dungi încrucișate este, de asemenea, împărțit în două tipuri principale:
- scheletice;
- cardiac.
Numele în sine reflectă principalele zone de localizare din corp. Funcțiile sale sunt extrem de importante, deoarece acest mușchi este cel care asigură contracția inimii, mișcarea membrelor și a tuturor celorlalte părți în mișcare ale corpului. Cu toate acestea, mușchii netezi nu sunt mai puțin semnificativi. Care sunt caracteristicile sale, vom analiza în continuare.
În general, se poate observa că doar munca coordonată efectuată de țesutul muscular neted și striat permite întregului organism să funcționeze cu succes. Prin urmare, este imposibil să se determine mai mult sau mai puțin semnificative dintre ele.
Caracteristici structurale netede
Principalele caracteristici neobișnuite ale structurii luate în considerare sunt structura și compoziția celulelor sale - miocite. Ca oricare altul, acest țesut este format dintr-un grup de celule care sunt similare ca structură, proprietăți, compoziție și funcții. Caracteristicile generale ale structurii pot fi identificate în mai multe puncte.
- Fiecare celulă este înconjurată de un plex dens de fibre de țesut conjunctiv care arată ca o capsulă.
- Fiecare unitate structurală se învecinează strâns cu cealaltă, spațiile intercelulare sunt practic absente. Acest lucru permite întregului material să fie strâns, structurat și puternic.
- Spre deosebire de colegul striat, această structură poate include celule de formă inegală.
Aceasta, desigur, nu este întreaga caracteristică că trăsăturile structurale, așa cum am menționat deja, se află tocmai în miocite, funcționarea și compoziția lor. Prin urmare, această problemă va fi discutată mai detaliat mai jos.

miocite musculare netede
Miocitele au forme diferite. În funcție de localizarea într-un anumit organ, acestea pot fi:
- oval;
- în formă de fus alungită;
- rotunjite;
- proces.
Cu toate acestea, în orice caz compozitia generala asemănătoare lor. Acestea conțin organite precum:
- mitocondrii bine definite și funcționale;
- complexul Golgi;
- miezul, adesea de formă alungită;
- reticul endoplasmatic;
- lizozomi.
Desigur, este prezentă și citoplasma cu incluziunile obișnuite. Un fapt interesant este că miocitele musculare netede sunt acoperite la exterior nu numai cu o membrană plasmatică, ci și cu o membrană (bazală). Acest lucru le oferă oportunitate suplimentară să se contacteze unul pe altul.
Aceste puncte de contact constituie caracteristicile țesutului muscular neted. Locurile de contact se numesc nexus-uri. Prin ele, precum și prin porii care se află în aceste locuri ale membranei, are loc transmiterea impulsurilor între celule, schimbul de informații, molecule de apă și alți compuși.
Există o altă caracteristică neobișnuită pe care o are țesutul muscular neted. Caracteristicile structurale ale miocitelor sale sunt că nu toate au terminații nervoase. De aceea, legăturile sunt atât de importante. Astfel încât nici o singură celulă nu rămâne fără inervație, iar impulsul poate fi transmis prin structura vecină prin țesut.
Există două tipuri principale de miocite.
- Secretar. Funcția lor principală este producerea și acumularea de granule de glicogen, conservarea multor mitocondrii, polizomi și unități ribozomale. Aceste structuri și-au primit numele datorită proteinelor conținute în ele. Acestea sunt filamente de actină și filamente de fibrină contractilă. Aceste celule sunt cel mai adesea localizate de-a lungul periferiei țesutului.
- Netede Acestea arată ca niște structuri alungite în formă de fus care conțin un nucleu oval, deplasat la mijlocul celulei. Un alt nume pentru leiomiocite. Ele diferă prin faptul că sunt mai mari. Unele particule din organul uterin ajung la 500 de microni! Aceasta este o cifră destul de semnificativă pe fundalul tuturor celorlalte celule din corp, cu excepția, poate, a oului.
Funcția miocitelor netede este, de asemenea, aceea de a sintetiza următorii compuși:
- glicoproteine;
- procolagen;
- elastan;
- substanță intercelulară;
- proteoglicani.
Interacțiunea comună și munca bine coordonată a tipurilor indicate de miocite, precum și organizarea acestora, asigură structura țesutului muscular neted.
Originea acestui mușchi
Există mai mult de o sursă de formare a acestui tip de mușchi în organism. Există trei origini principale. Așa se explică diferențele pe care le are structura țesutului muscular neted.
- origine mezenchimală. majoritatea fibrelor netede au acest lucru. Din mezenchim se formează aproape toate țesuturile care căptușesc interiorul organelor goale.
- origine epidermica. Numele în sine vorbește despre locurile de localizare - acestea sunt toate glandele pielii și canalele lor. Acestea sunt formate din fibre netede care au această variantă de aspect. Transpirație, salivare, lapte, lacrimal - toate aceste glande își secretă secretul din cauza iritației celulelor mioepiteliocitelor - particulele structurale ale organului în cauză.
- origine neuronală. Astfel de fibre sunt localizate într-un loc specific - acesta este irisul, una dintre membranele ochiului. Contracția sau expansiunea pupilei este inervată și controlată de aceste celule musculare netede.
În ciuda originilor diferite, compoziția internă și proprietățile de performanță ale tuturor țesuturilor luate în considerare rămân aproximativ aceleași.
Principalele proprietăți ale acestei țesături
Proprietățile țesutului muscular neted corespund cu cele ale țesutului muscular striat. În aceasta sunt uniți. Acest:
- conductivitate;
- excitabilitate;
- labilitate;
- contractilitatea.
În același timp, există o caracteristică destul de specifică. Dacă mușchii scheletici striați sunt capabili să se contracte rapid (aceasta este o bună ilustrare a tremurului în corpul uman), atunci cel neted poate fi ținut în stare comprimată pentru o lungă perioadă de timp. În plus, activitățile sale nu sunt supuse voinței și minții omului. Pentru că o inervează
O proprietate foarte importantă este capacitatea de a se întinde pe termen lung (contracție) și aceeași relaxare. Deci, aceasta este baza activității vezicii urinare. Sub influența fluidului biologic (umplerea acestuia), este capabil să se întindă și apoi să se contracte. Pereții săi sunt căptușiți cu mușchi netezi.

Proteinele celulare
Miocitele țesutului în cauză conțin multe compuși diferiți. Cu toate acestea, cele mai importante dintre ele, care asigură funcțiile de contracție și relaxare, sunt tocmai moleculele proteice. Dintre acestea, iată:
- filamente de miozină;
- actină;
- nebulină;
- conectarea;
- tropomiozina.
Aceste componente sunt de obicei localizate în citoplasma celulelor izolate unele de altele, fără a forma clustere. Cu toate acestea, în unele organe la animale se formează mănunchiuri sau fire numite miofibrile.
Locația în țesutul acestor mănunchiuri este în principal longitudinală. Mai mult, atât fibrele de miozină, cât și fibrele de actină. Ca urmare, se formează o întreagă rețea în care capetele unora sunt împletite cu marginile altor molecule de proteine. Acest lucru este important pentru contracția rapidă și corectă a întregului țesut.
Contracția în sine are loc astfel: în compoziția mediului intern al celulei există vezicule pinocitare, care conțin în mod necesar ioni de calciu. Când ajunge impuls nervos, indicând necesitatea contracției, această bulă se apropie de fibrilă. Ca urmare, ionul de calciu irită actina și se deplasează mai adânc între filamentele de miozină. Aceasta duce la implicarea plasmalemei și ca urmare, miocitul este redus.

Țesutul muscular neted: desen
Dacă vorbim despre țesut striat, atunci este ușor să-l recunoaștem după striația sa. Dar în ceea ce privește structura pe care o luăm în considerare, acest lucru nu se întâmplă. De ce țesutul muscular neted are un model complet diferit față de vecinul său apropiat? Acest lucru se datorează prezenței și locației componentelor proteice în miocite. În compoziția mușchilor netezi, filamentele miofibrilelor de natură diferită sunt localizate haotic, fără o stare ordonată definită.
De aceea modelul țesăturii este pur și simplu absent. În filamentul striat, actina este înlocuită succesiv cu miozină transversală. Ca rezultat, apare un model - striație, datorită căruia țesătura și-a primit numele.
La microscop, țesutul neted arată foarte uniform și ordonat, datorită miocitelor alungite situate longitudinal, strâns adiacente unele cu altele.
Zone de aranjare spațială în corp
Țesutul muscular neted produce suficient un numar mare de organe interne importante în corpul animalului. Deci, ea a fost educată:
- intestine;
- organele genitale;
- vase de sânge de toate tipurile;
- glande;
- organele sistemului excretor;
- Căile aeriene;
- părți ale analizorului vizual;
- organele sistemului digestiv.
Evident, locurile de localizare ale țesutului în cauză sunt extrem de diverse și importante. În plus, trebuie remarcat faptul că astfel de mușchi formează în principal acele organe care sunt supuse controlului automat.

Metode de recuperare
Țesutul muscular neted formează structuri care sunt suficient de importante pentru a avea capacitatea de a se regenera. Prin urmare, se caracterizează prin două modalități principale de recuperare a daunelor de diferite tipuri.
- Diviziunea mitotică a miocitelor până când se formează cantitatea necesară de țesut. Cel mai comun mod simplu și rapid de regenerare. Așa are loc refacerea părții interne a oricărui organ format din mușchii netezi.
- Miofibroblastele sunt capabile să se transforme în miocite țesătură netedă daca este necesar. Aceasta este o modalitate mai complexă și mai rară de regenerare a acestui țesut.
Inervația mușchilor netezi
Țesutul muscular neted își îndeplinește funcțiile indiferent de dorința sau nedorința unei ființe vii. Acest lucru se datorează faptului că inervația sa este efectuată de vegetativ sistem nervos, precum și procesele nervilor ganglionilor (coloana vertebrală).
Un exemplu în acest sens și dovadă este reducerea sau creșterea dimensiunii stomacului, ficatului, splinei, întinderea și contracția vezicii urinare.

Funcțiile țesutului muscular neted
Care este sensul acestei structuri? De ce aveți nevoie de netezire, următoarele:
- contracția prelungită a pereților organelor;
- dezvoltarea secretelor;
- capacitatea de a răspunde la stimuli și expunere cu excitabilitate.













